In traditional view, Viruses, with their often small genomes and error-prone replication mechanisms, possess extraordinary adaptive abilities and can display rates of sequence change that are orders of magnitude greater than those of the hosts they infect, viruses can quickly adapt to new hosts and evade innate and adaptive immune responses. It is precisely because of these abilities that they may also help to create the evolutionary stasis of viruses in long-term host relationships. Extreme genetic conservation of viruses over longer periods of evolution, virus evolution conditioned by hosts they infect, host represents the primary driver of the longer-term evolution of viruses.
A large body of literature documents remarkably high nucleotide substitution rates in virus genomes, virus sequence change occurs so quickly that phylogenetic trees of their genes are often temporally structured-viruses from older samples show systematically less divergence from the most recent common ancestor (MRCA) than those collected more recently. Furthermore, virus evolution has often been linked to historical events. Virus sequence change is often dominated by synonymous substitutions in coding regions that leave sequences of the encoded proteins unaltered. Fixation of these changes may be facilitated by repeated transmission bottlenecks that reduce effective population sizes that occur as the virus transmits between hosts. Sequence change may be augmented by adaptive changes, influenza A virus, HIV-1 and HCV are typical examples. These observations contribute to a general perception of the ephemeral nature of RNA viruses and a broader idea that viruses are rapidly evolving entities with perhaps frequent recent origins.
Host-driven virus evolution
Host-driven viral evolution can be explained by niche-filling models that emphasize the role of host interactions in shaping virus evolution. As viruses can rapidly evolve to a fitness peak in a given host environment, this may have the paradoxical effect of restricting sequence change rather than accelerating it in any period other than the short term. Infection of the same host over tens or hundreds of years or perhaps even millennia may drive the evolution of each host-adapted virus to evolutionary stasis-an optimized genome that is maximized in those aspects of its fitness that maintain infections in the host population. In the niche-filling models, once fully adapted to their niche, the intensity of peer competition may create virus genomes with few genuinely phenotypically neutral sites.
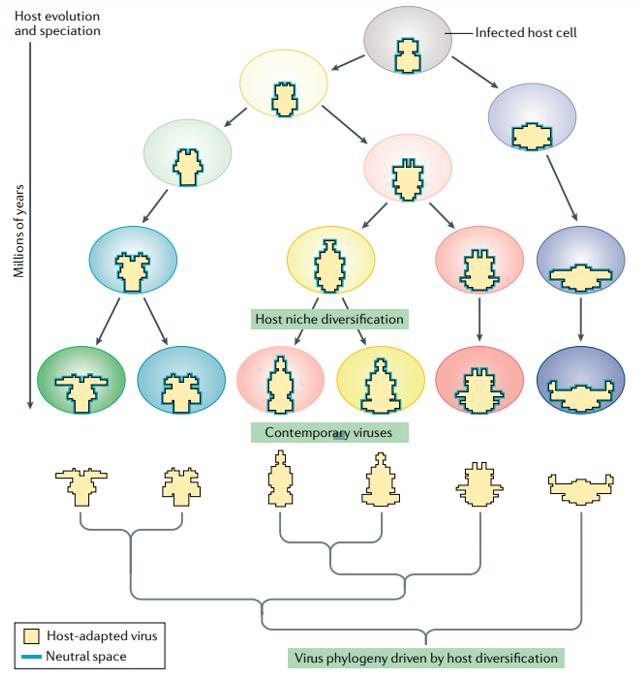 Fig. 1 Host-driven virus evolution.
Fig. 1 Host-driven virus evolution.
(Source: Nat Rev Microbiol. May 2019.)
The process of host adaptation generates viruses that are primarily shaped by the constraints of the niche and less by the ancestry of the virus. virus genome sequence change driven by host change resembles niche-filling models developed for phenotypic trait evolution in cellular organisms, traits evolve adaptively to fit the niche. The niche is defined by the host organism that the virus infects, the viral sequence defines the phenotype, and changes are primarily adaptive. These changes ultimately drive longer-term evolution of viruses and explain why their nucleotide substitution rates ultimately approach those of their hosts.
Host jumps can be viewed as the occupancy of a new niche and hence a rapid adaptation of trait values to fit this niche. Host jumps are associated with periods of accelerated sequence change as the virus remodels and regains fitness in an altered environment. Host adaptation after cross-species transmission is associated with rapid amino acid sequence changes of viral genes, typically those associated with receptor interactions and the evasion of innate immunity but often pervasive throughout the entire virus genome.