In vivo gene therapy data using AAV as a vector, especially clinical trial data, suggest that the immune response to AAV vectors is not strong when administered at low doses, but may cause a relatively strong inflammatory immune response,affecting tissues and organs, and produce toxic effects when administered at high doses. In addition, immune responses to AAV vectors and their therapeutic gene products can inhibit their efficacy. Therefore, it is necessary to conduct in-depth research on the immune response to AAV infection, including innate immunity and adaptive immune response, to guide the best drug strategy or program, on the one hand, to ensure the safety of AAV gene therapy, and on the other hand, to improve the efficacy of AAV gene drugs effect.
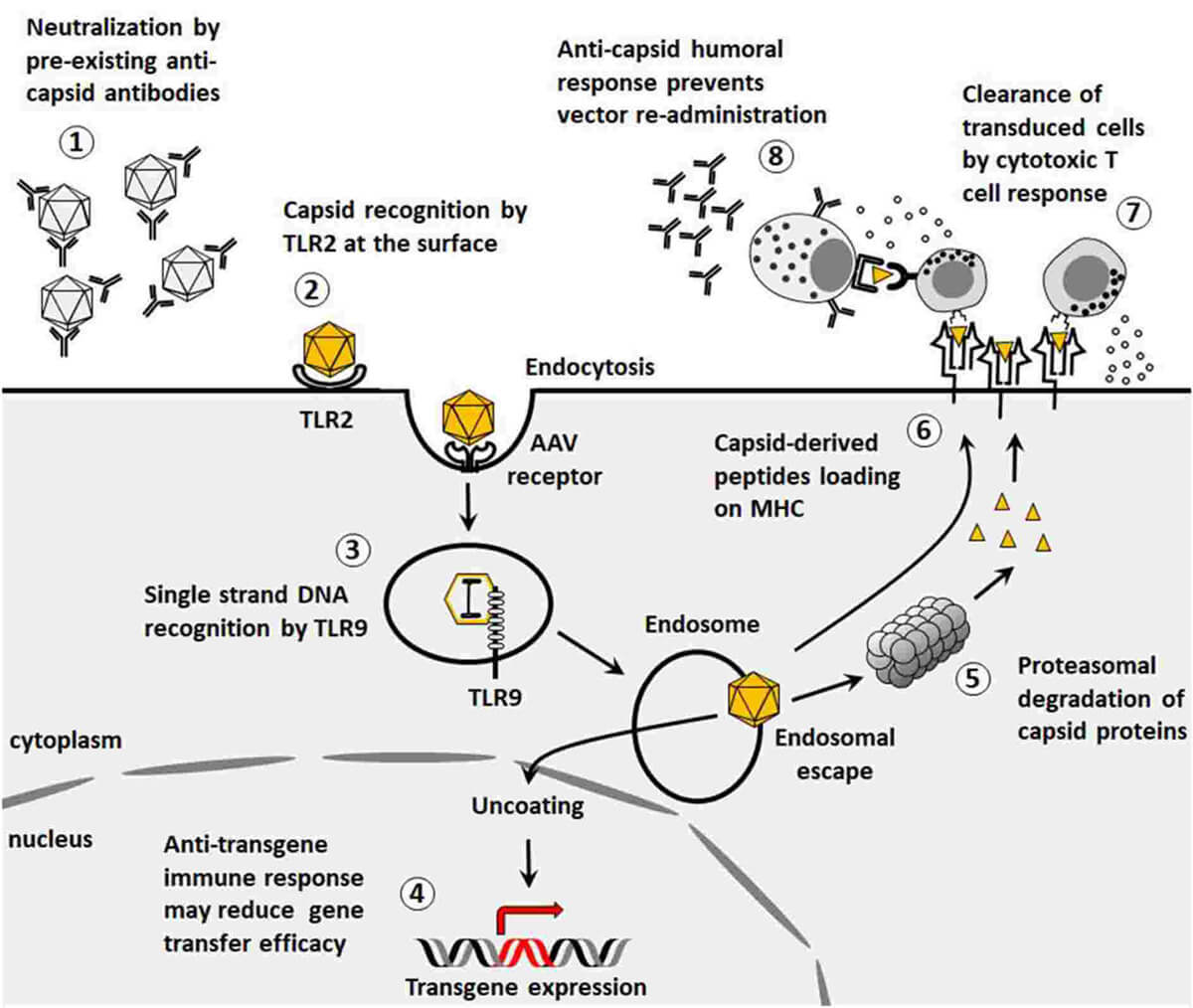 Figure 1. Immunological barriers to gene transfer. (Ronzitti G, et al.; 2020)
Figure 1. Immunological barriers to gene transfer. (Ronzitti G, et al.; 2020)
When the AAV vector enters human cells, the antigen-presenting cells (such as DCs, macrophages, etc.) in the body recognize the pathogen-related molecular patterns on the AAV vector through pattern recognition receptors (such as TLR9, TLR2, etc.) and activate downstream related processes signaling pathway, activating NF-κB (nuclear factor kB), IRF (IFN-regulatory factor) and other transcription factors, which play a central role in inducing pro-inflammatory factors and type I IFN respectively. The cytokines induced by the activation of the above signaling pathways can Stimulates the maturation of antigen-presenting cells and upregulates the expression of MHC and some costimulatory factors. When the antigen-presenting cells migrate to lymph nodes, they present the AAV vector antigen peptide to lymphocytes, thereby activating sensitized antigen-specific T cells and specific immune responses.
Toll-like receptors (TLRs) are a type of transmembrane receptors that participate in body immunity. Eleven members of the Toll-like receptor family have been discovered in mammals. Among them, the more clearly studied ones are TLR2, TLR4, TLR5 and TLR9. TLRs are like the eyes of innate immunity, monitoring and recognizing various pathogen-associated molecular patterns (PAMPs). Some TLR family members can not only recognize foreign substances such as exogenous pathogens, but also recognize endogenous substances and degradation products. TLR9 is a pattern recognition receptor that recognizes CpGDNA. It can activate NF-κB and AP-1 by activating signal transduction in the monocyte/phagocytic cell system, thereby releasing a large amount of TNF-α, IL-1, IL-6 and IL. 12 and other pro-inflammatory cytokines, causing acute inflammatory response, sepsis and even shock. TLR2 is a Toll-like receptor with the largest recognition range of ligands in the Toll-like family. It can recognize a variety of ligands including lipoproteins, lipopolypeptides, lipoteichoic acid (LTA), arabinomannan (LAM), and zymosan. Activated TLR2 can play an important role in the immune system and trigger specific immune responses. For TLR recognition of ligands, different types of TLRs can be combined to recognize different PAMPs. For example, TLR7 may be combined with TLR9 to mediate CpG activation of immune cells. Among them, TLR4/TLR4 and TLR9/TLR9 are in the form of homodimers; while TLR2/TLR4, TLR2/TLR6 and TLR7/TLR8 are heterodimers, and some dimers have a subunit that has not yet been OK, such as TLR3/TLR, TLR5/TLR.
TLR9 in cells such as plasmacytoid DCs (pDCs) in the blood and Kupffer cells in the liver is the main pattern recognition receptor that recognizes AAV genomic nucleic acids (especially unmethylated CpG DNA). Some researchers have used single-stranded AAV vector stimulated mouse-derived pDC in vitro, and an increase in type I IFN was detected in the culture supernatant. Similar results were obtained using human peripheral mononuclear cells (PBMC). Preclinical studies have also confirmed that activation of the TLR9-MyD88 signaling pathway is one of the causes of immune activation in AAV gene therapy targeting liver and muscle. Single-chain AAV vectors stimulated pDCs from TLR9 knockout mice and did not show an increase in IFN secretion. After treating human pDCs with TLR9 inhibitors, stimulation with single-chain AAV was unable to increase IFN secretion. In addition, studies have shown that the effect of AAV vector on stimulating pDCs to secrete IFN has nothing to do with the AAV vector serotype and the type of genetic element loaded. When single-chain AAV vectors were used to stimulate mouse macrophages, Kupffer cells, and human monocytes, no increase in the secretion of type I IFN was found. The above results indicate that TLR9 is a pathogen pattern recognition receptor in pDC that recognizes AAV vectors. pDC is the most important cell in the body for producing type I IFN, and its efficiency in producing type I IFN is more than a hundred times that of other cells. Researchers have found that type I IFN is an important factor in inducing anti-AAV capsid CD8+ T cell responses in specific immunity. Innate immune cells pDC and cDC work together to activate CD8+ T cell immune responses in specific immune responses, among which TLR9 is pDC is necessary. For cDC, TLR9 is not important. pDC monitors the invasion of AAV vector through TLR9, and cDC is responsible for the presentation of AAV capsid polypeptide antigen. TLR9 knockout mice can restore their CD8+ T cell immune responses against AAV capsids by adoptive infusion of wild-type mouse pDCs. The above studies indicate that the TLR9 innate immune signaling pathway in pDC is an important factor in subsequent adaptive cellular immune CD8+ T activation.
Initial T cells mature in the thymus, then migrate out of the thymus, enter the blood circulation, home and settle in peripheral lymphoid organs, and recirculate in the body. Initial T cells specifically bind to the AAV antigen-MHC molecule complex (pMHC) on the surface of APC through their TCR, activate and proliferate under the combined action of costimulatory signals and cytokines, and then differentiate into effector T cells to complete the clearance of the antigen and modulation of immune responses. T cell-mediated immune response, also called cellular immune response, is a continuous process that can be divided into three stages: T cell specific antigen recognition stage; T cell activation, proliferation and differentiation stage; Generation and effector stage of effector T cells.
From the above, it can be seen that AAV activates adaptive cellular immune CD8+ T cells in pDC through the TLR9 innate immune signaling pathway. This triggers the activation, proliferation and differentiation of T cells, producing T cell types such as Th1, Th2, Th17, and Treg. Among them, Th1 can induce CTL differentiation and release of cytokines through direct contact to recruit and activate monocytes/macrophages and lymphocytes to induce cellular immune responses; Th2 can assist B cell activation through direct contact, assisting and promoting the proliferation and differentiation of B cells. They are plasma cells that produce antibodies to neutralize AAV; Th17 exerts effects by secreting cytokines; Treg cells can play a negative immune regulatory role through a variety of mechanisms. Usually, the body's immune response and immune effects to specific antigens do not last long. Once the AAV antigens have been cleared, the immune system must return to balance. Therefore, effector cells also need to be inhibited or eliminated, leaving only a few memory cells to maintain immune memory so that a rapid response can occur when exposed to the antigen again.
Anti-AAV Antibody ELISA Kit
AAV Antibodies and Titration ELISA
References
| Target | Cat. No. | Product Name | Size | Species Reactivity | Application | Detection Sample | |
| AAV1 | DEIASL342 | AAV9 Titration ELISA Kit | 96T | Qualitative | serum, plasma | Inquiry | |
| AAV2 | DEIASL347 | AAV6 Titration ELISA Kit | 96T | Human | Qualitative | Serum | Inquiry |
| DEIASL347M | Mouse Anti-AAV8 ELISA Kit(Quantitative) | 96T | Mouse | Qualitative | Serum | Inquiry | |
| AAV5 | DEIASL343 | AAV8 Titration ELISA Kit | 96T | Qualitative | serum, plasma | Inquiry | |
| AAV6 | DEIAAV6 | Anti-AAV2 antibody ELISA Kit | 96T | N/A | Quantitative | cell culture supernatants, purified virus preparations | Inquiry |
| DEIASL344 | Anti-AAV9 antibody ELISA Kit | 96T | Human | Qualitative | Serum | Inquiry | |
| DEIASL344M | Anti-AAV1 Antibody ELISA Kit | 96T | Mouse | Qualitative | Serum | Inquiry | |
| AAV8 | DEIASL345MQ | Anti-AAV5 Antibody ELISA Kit | 96T | Mouse | Quantitative | Serum | Inquiry |
| DEIAAV8 | Anti-AAV6 ELISA Kit | 96T | N/A | Quantitative | cell culture supernatants, purified virus preparations | Inquiry | |
| DEIASL345 | Anti-AAV8 antibody ELISA Kit | 96T | Human | Qualitative | Serum | Inquiry | |
| DEIASL345M | Mouse Anti-AAV9 antibody ELISA Kit | 96T | Mouse | Qualitative | Serum | Inquiry | |
| AAV9 | DEIAAV9 | Mouse Anti-AAV2 antibody ELISA Kit | 96T | N/A | Quantitative | cell culture supernatants, purified virus preparations | Inquiry |
| DEIASL348 | Mouse Anti-AAV6 antibody ELISA Kit | 96T | Human | Qualitative | Serum | Inquiry | |
| DEIASL348M | Mouse Anti-AAV8 antibody ELISA Kit | 96T | Mouse | Qualitative | Serum | Inquiry |


