Adeno-associated virus is a single-stranded DNA virus, and the current scientific consensus is that it does not cause any human disease. It consists of a protein capside and a single-stranded DNA genome of 4.7kb in length. The protein capsid is composed of three subunits, namely VP1, VP2, and VP3. The two ends of the AAV genome are two "T"-shaped inverted terminal repeats (ITR). These two ITRs are the origin of viral DNA replication and signals that trigger viral packaging. The rep gene in the AAV genome encodes four proteins related to viral replication, namely Rep78, Rep68, Rep52, and Rep40.
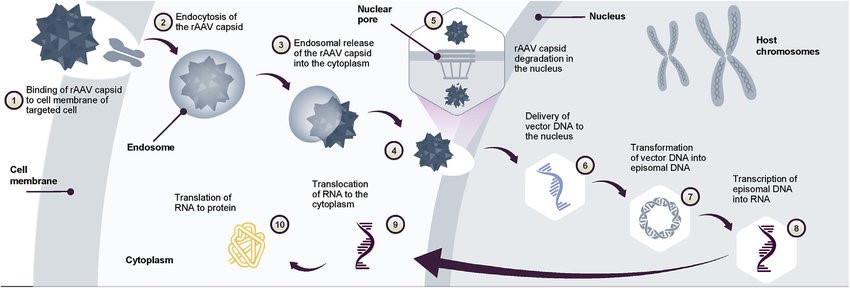 Figure 1. AAV gene transfer therapy mechanism of action Following gene transfer therapy administration.(Mendell JR, et al.; 2022)
Figure 1. AAV gene transfer therapy mechanism of action Following gene transfer therapy administration.(Mendell JR, et al.; 2022)
The protein capsid carried by the recombinant adeno-associated virus (rAAV) used as a gene therapy vector is almost identical to that of wild-type AAV. However, the portion of the genome within the capsid that encodes the viral protein is completely deleted and replaced by a therapeutic transgene. The only retained part of the AAV genome is the ITRs, which serve to guide genome replication and viral vector assembly. The advantage of completely deleting the part encoding the viral protein is that on the one hand, it can maximize the capacity of the recombinant AAV to carry the transgene, and on the other hand, it can reduce the immunogenicity and cytotoxicity produced when the transgene is delivered in vivo.
Recombinant AAV particles enter cells through clathrin-mediated endocytosis by binding to glycosylation receptors expressed on the host cell surface. After acidification of the endosome formed by endocytosis, the conformation of the VP1/VP2 part of the viral capsid changes, causing the virus to escape from the endosome and enter the nucleus through the nuclear pore. After entering the nucleus, the single-stranded DNA is released from the capsid. At this point, single-stranded DNA cannot yet be transcribed, and they need to become double-stranded DNA. Single-stranded DNA can be synthesized using the host cell's DNA polymerase to synthesize complementary strands, or two complementary strands released from different AAV particles can anneal to form double-stranded DNA.
The latest AAV genome design can design the single-stranded DNA carried in the capsid into a self-complementary sequence. The advantage of this sequence is that it can be transcribed without the step of replicating single-stranded DNA into double-stranded DNA. Compared with the traditional single-stranded AAV genome, its gene expression is faster and the expression level is higher. One drawback of this design, however, is that the transgene capacity carried by the AAV is halved.
The double-stranded AAV genome then uses ITRs to undergo intramolecular or intermolecular genome recombination. This process allows the AAV genome to become stable episomal DNA, resulting in the genome being able to continue gene expression in cells that are no longer undergoing mitosis.
The tissues targeted by most gene therapy development programs are the liver, striated muscle, and the central nervous system. Almost all native AAV capsid proteins are capable of triggering efficient transgene expression in the liver, so liver-targeted recombinant AAVs provide an excellent gene delivery platform for the treatment of a range of diseases. They include diseases such as hemophilia A and B and familial hypercholesterolemia.
The ability of AAV8 and AAV9 capsid proteins to target multiple muscle types in the body allows AAV-mediated gene therapy to be used to treat a variety of muscle diseases, including Duchenne muscular dystrophy (DMD). It is worth mentioning that muscles can serve as "factories in the body" for the production of therapeutic molecules, so gene therapy targeting muscle tissue can be used to treat non-muscle diseases.
Another important direction for recombinant AAV-delivered gene therapy is the CNS, including the eyes and brain. The eye is a relatively isolated environment. Direct intraocular injection and delivery of AAV gene therapy can achieve the effect of treating a variety of genetic eye diseases.
Gene replacement: The goal of this strategy is to deliver genes that express normal proteins to compensate for the effects of loss-of-function mutations. It is suitable for treating recessive single-gene diseases and has achieved great success in clinical trials. Glybera and Luxturna are good examples. Glybera is based on a recombinant AAV1 platform that delivers a transgene encoding lipoprotein lipase (LPL) and is used to treat patients with LPL deficiency. Luxturna uses the AAV2 platform to deliver the normal RPE65 gene. Both treatments use local injections into the patient's muscles and eyes.
The discovery of newer AAV serotypes allows researchers to deliver transgenes throughout the body through intravenous injection of AAV vectors. These new AAV vectors bring effective therapies for the treatment of hemophilia A and B (targeting the liver), DMD (targeting muscles throughout the body), and SMA (targeting the CNS, including the spinal cord). One example is Zolgensma, a gene therapy that uses an AAV9 vector to deliver an SMA transgene encoding the survival motor neuron protein (SMN). In many diseases, simply expressing normal genes in some cells is enough to alleviate symptoms.
Gene silencing: As opposed to gene replacement, the goal of gene silencing is to treat single-gene diseases caused by gain-of-function gene mutations, such as Huntington disease. RNAi therapy is currently the preferred strategy for AAV gene silencing platforms. However, compared with the rapid progress of synthetic RNAi therapies, RNAi therapies based on AAV delivery platforms are still in the preclinical development stage. Gene silencing needs to occur in most tissues to have a meaningful therapeutic effect. In some organs (such as the brain), this is still a challenge for AAV vectors.
In addition to the RNAi strategy, using the CRISPR-Cas system to target RNA is also a gene silencing strategy. However, the obstacles that this strategy needs to overcome are the limited size of the transgene that recombinant AAV can accommodate, and the Cas protein found in bacteria may trigger an immune response, thus reducing the efficacy.
Delivery of new genes: In addition to treating single-gene diseases, AAV-mediated gene therapy has the potential to treat complex genetic diseases or infectious diseases by delivering new genes. For example, heart failure and infectious diseases represent disease areas with high unmet need, and by introducing novel genes, gene therapy can deliver growth factors, modulate signaling pathways associated with heart failure, and express antibodies that neutralize lethal viral infections. Current clinical trials are testing treatments for HIV infection that use muscle as a "biofactory" to produce therapeutic antibodies.
Gene therapy is an innovative treatment platform that has gained industry attention. As a powerful means of delivering transgenes, the research and development of AAV vector platforms is also experiencing exponential growth. Although the study of AAV belongs to the field of virology, the process of converting AAV into a gene therapy vector requires multidisciplinary joint research. Breakthroughs in molecular biology, bioinformatics, infectious diseases, structural biology, immunology, and genomics all play an indispensable role in the successful development of AAV vector platforms.
The development of AAV vector platforms still needs to overcome several challenges, including how to cost-effectively generate sufficient quantities of therapeutic AAV vectors and overcoming the immune rejection of AAV vectors and transgenic products by the human immune system. Overcoming these challenges is critical to the promotion of AAV-mediated gene therapy.
Anti-AAV Antibody ELISA Kit
AAV Antibodies and Titration ELISA
Reference
| Target | Cat. No. | Product Name | Size | Species Reactivity | Application | Detection Sample | |
| AAV1 | DEIASL342 | AAV9 Titration ELISA Kit | 96T | Qualitative | serum, plasma | Inquiry | |
| AAV2 | DEIASL347 | AAV6 Titration ELISA Kit | 96T | Human | Qualitative | Serum | Inquiry |
| DEIASL347M | Mouse Anti-AAV8 ELISA Kit(Quantitative) | 96T | Mouse | Qualitative | Serum | Inquiry | |
| AAV5 | DEIASL343 | AAV8 Titration ELISA Kit | 96T | Qualitative | serum, plasma | Inquiry | |
| AAV6 | DEIAAV6 | Anti-AAV2 antibody ELISA Kit | 96T | N/A | Quantitative | cell culture supernatants, purified virus preparations | Inquiry |
| DEIASL344 | Anti-AAV9 antibody ELISA Kit | 96T | Human | Qualitative | Serum | Inquiry | |
| DEIASL344M | Anti-AAV1 Antibody ELISA Kit | 96T | Mouse | Qualitative | Serum | Inquiry | |
| AAV8 | DEIASL345MQ | Anti-AAV5 Antibody ELISA Kit | 96T | Mouse | Quantitative | Serum | Inquiry |
| DEIAAV8 | Anti-AAV6 ELISA Kit | 96T | N/A | Quantitative | cell culture supernatants, purified virus preparations | Inquiry | |
| DEIASL345 | Anti-AAV8 antibody ELISA Kit | 96T | Human | Qualitative | Serum | Inquiry | |
| DEIASL345M | Mouse Anti-AAV9 antibody ELISA Kit | 96T | Mouse | Qualitative | Serum | Inquiry | |
| AAV9 | DEIAAV9 | Mouse Anti-AAV2 antibody ELISA Kit | 96T | N/A | Quantitative | cell culture supernatants, purified virus preparations | Inquiry |
| DEIASL348 | Mouse Anti-AAV6 antibody ELISA Kit | 96T | Human | Qualitative | Serum | Inquiry | |
| DEIASL348M | Mouse Anti-AAV8 antibody ELISA Kit | 96T | Mouse | Qualitative | Serum | Inquiry |


