AAV is a single-stranded linear DNA defective virus with a genomic DNA less than 5 kb, and its structure is an icosahedral non-enveloped particle. AAV consists of a single-stranded DNA with an anti-terminal repeat sequence and two open reading frames Rep and Cap at both ends. IRT is a symmetric repeat sequence that plays an important role in the structure and function of AAV. The Rep gene consists of four overlapping genes, Rep78, Rep68, Rep52, and Rep40, which encode the Rep protein required for AAV replication, packaging, and genome integration. Cap encodes capsid proteins VP1, VP2 and VP3, and the ratio of capsid proteins is 1:1:10 (VP1:VP2:VP3). These three molecules interact to form a symmetrical icosahedral structure that serves as a carrier for gene delivery.
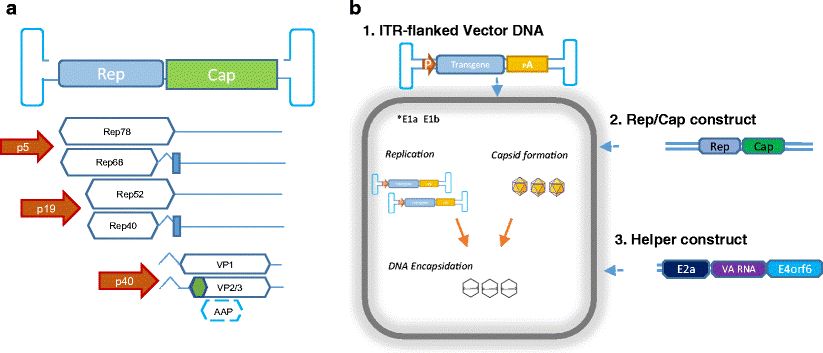 Figure 1. Adeno-associated virus (AAV) vector biology. (Aponte-Ubillus JJ, et al.; 2018)
Figure 1. Adeno-associated virus (AAV) vector biology. (Aponte-Ubillus JJ, et al.; 2018)
The AAV genome is a linear, single-stranded (ssDNA) molecule containing 4680 nucleotides and a 145-base terminal repeat sequence (ITR) at its end. The ITR sequence folds into a hairpin structure and serves as the only known cis -acting element required for the initiation of DNA replication and packaging of recombinant AAV genomes into infectious virions. The ITR element is located on either side of two translated open reading frames (ORFs). The left ORF (or rep gene) encodes four nonstructural Rep proteins, which are critical for viral DNA replication and packaging. There are 2 promoters on the left side of the AAV genome, namely p5 and p19, which can produce 2 different lengths of mRNA, each containing an alternatively spliced intron, so 4 different mRNAs containing overlapping sequences can be produced. The Rep proteins, namely Rep78, Rep68, Rep52, and Rep40, encode the Rep proteins required for AAV replication, packaging, and genome integration. Rep78 and Rep68 proteins can specifically bind to the hairpin structure formed by ITR and cleave at specific sites. All Rep proteins have ATP-binding activity and helicase activity, which can up-regulate the transcription of p40 promoter and down-regulate the activities of p5 and p19 promoters. The ORF (or cap gene) on the right encodes three structural proteins, VP1, VP2, and VP3. VP1 is translated from a 2.6 kb mRNA, while VP2 and VP3 are translated from a 2.3 kb mRNA. The start codons of VP1, VP2 and VP3 are AUG (2 203), ACG(2 614) and AUG(2 809) respectively. ACG(2 614) is a start codon unique to eukaryotic cells. Its Starting capability is lower than AUG. If ACG (2 614) in the cap gene is mutated to AUG (2 614), VP2 expression can be increased, but VP3 expression will be reduced.
In addition to participating in the replication and packaging of AAV, the function of Rep protein is mainly manifested in the site-specific integration of the AAV genome. The necessary elements for AAV site-directed integration include: ITRs (cis-acting element), Rep78 or Rep68 protein (trans-acting factor) and a 138 bp integration efficiency element (IEE) inside the P5 promoter. Whether this IEE element within the P5 promoter is necessary awaits further study, as a recent study suggested that a 16 bp sequence in the RBE (Rep binding element) of the P5 promoter is sufficient. Tissue culture experiments indicate that latent infection with wild-type AAV2 is non-pathogenic when the Rep protein is expressed under the control of its own promoter. The expression of Rep protein is regulated by negative feedback. Excessive Rep protein expression inhibits cell division and induces programmed cell death.
There is a 33 bp sequence in the AAVS1 site, which contains an RBE-like and a TRS (Terminal resolution site)-like sequence, which are separated by 8 nucleotides. This sequence is necessary for AAV integration. The integration sequences can be different, but the 5'CTC must be present in all. The actual integration site is downstream of the sequence of interest and is variable. There are many RBE sequences in the human genome, but AAVS1 is the only site in close proximity to an RBE and a TRS. In vitro experiments show that the AAV genome and AAVS1 can be entangled with each other through the action of Rep68 protein. These experiments can only explain at the molecular level why the AAVS1 site is localized, but the specific mechanism remains unknown.
Even in the case of ideal expression of Rep78/68 protein, site-directed insertion is not completely specific, and about 40% to 70% of insertions occur at the AAVS1 site. Furthermore, the insertion was not very precise because there were no repetitive breakpoints in the vector-AAVS1 junction; however, the integrated cluster emerged as a 2 kb fragment on AAVS1. Recently, a study showed that an intracellular protein, TAR-RNA loop binding protein 185 (TRP-185), can integrate AAV into the AAVS1 site further downstream of the RBE by interacting with the Rep protein and the AAVS1 site. Furthermore, transgene integration into the AAVS1 locus in mice and rats demonstrated the presence of an AAVS1-open chromatin structure in vivo and the presence of intracellular factors mediating site-specific integration in nondividing cells.
Studies have found that the non-structural protein Rep78 of AAV2 can inhibit tumor transformation, with three main mechanisms: a. Promoting the function of tumor suppressor genes. At the protein level, Rep78 can interact with E2F-1 and stabilize the pRb-E2F-1 complex; At the DNA level, Rep78 can bind to the E2F-1 promoter and down-regulate the adenovirus-mediated transcription of E2F-1. This dual role of Rep protein reduces the free state of E2F-1 in the cell, thereby making it have the ability to suppress tumors. b. inhibit the expression of proto-oncogenes. Rep78 protein can down-regulate the expression of human c-fos and c-myc proto-oncogene promoters. c. Inhibiting viral oncogenes, Rep78 protein can inhibit the activity of the P97 promoter of HPV-16, thereby inhibiting the occurrence of cervical cancer. Other studies have found that overexpression of Rep78 protein can inhibit DNA synthesis in S phase. Rep78 can completely arrest cells in S phase, which rarely occurs after cellular DNA damage. The Rep78 protein inhibits the activity of Cdc25A by binding to two proteins that stabilize cell cycle regulatory phosphatase (Cdc25A), making it unable to activate the downstream substrates-cyclin-dependent kinases Cdk1 and Cdk2, while Cdk1 and Cdk2 are required during DNA replication. But this alone cannot completely arrest cells in S phase. The Rep78 protein nicks the cell's chromosomes, thereby introducing a cell cycle checkpoint kinase (ATM)-mediated DNA damage response. Activated ATM regulates the cell cycle by phosphorylating its downstream corresponding protein Chk2, ultimately causing the cell cycle to stay in the G1/G2 phase. Mutation analysis demonstrates that the zinc finger structure and nuclease activity of Rep78 are required for S phase arrest. The above factors work together to completely inhibit host DNA synthesis.
In short, compared with other viral vectors, AAV has unique advantages, which makes it have broad prospects in gene therapy, tumor prevention and treatment, etc. AAV itself also has a specific killing effect on some special tumor cells. By producing Rep protein, it interferes with the cell cycle and proliferation, and initiates programmed apoptosis of tumor cells. At the same time, it does not have any adverse effects on normal cells.
Anti-AAV Antibody ELISA Kit
AAV Antibodies and Titration ELISA
References
| Target | Cat. No. | Product Name | Size | Species Reactivity | Application | Detection Sample | |
| AAV1 | DEIASL342 | AAV9 Titration ELISA Kit | 96T | Qualitative | serum, plasma | Inquiry | |
| AAV2 | DEIASL347 | AAV6 Titration ELISA Kit | 96T | Human | Qualitative | Serum | Inquiry |
| DEIASL347M | Mouse Anti-AAV8 ELISA Kit(Quantitative) | 96T | Mouse | Qualitative | Serum | Inquiry | |
| AAV5 | DEIASL343 | AAV8 Titration ELISA Kit | 96T | Qualitative | serum, plasma | Inquiry | |
| AAV6 | DEIAAV6 | Anti-AAV2 antibody ELISA Kit | 96T | N/A | Quantitative | cell culture supernatants, purified virus preparations | Inquiry |
| DEIASL344 | Anti-AAV9 antibody ELISA Kit | 96T | Human | Qualitative | Serum | Inquiry | |
| DEIASL344M | Anti-AAV1 Antibody ELISA Kit | 96T | Mouse | Qualitative | Serum | Inquiry | |
| AAV8 | DEIASL345MQ | Anti-AAV5 Antibody ELISA Kit | 96T | Mouse | Quantitative | Serum | Inquiry |
| DEIAAV8 | Anti-AAV6 ELISA Kit | 96T | N/A | Quantitative | cell culture supernatants, purified virus preparations | Inquiry | |
| DEIASL345 | Anti-AAV8 antibody ELISA Kit | 96T | Human | Qualitative | Serum | Inquiry | |
| DEIASL345M | Mouse Anti-AAV9 antibody ELISA Kit | 96T | Mouse | Qualitative | Serum | Inquiry | |
| AAV9 | DEIAAV9 | Mouse Anti-AAV2 antibody ELISA Kit | 96T | N/A | Quantitative | cell culture supernatants, purified virus preparations | Inquiry |
| DEIASL348 | Mouse Anti-AAV6 antibody ELISA Kit | 96T | Human | Qualitative | Serum | Inquiry | |
| DEIASL348M | Mouse Anti-AAV8 antibody ELISA Kit | 96T | Mouse | Qualitative | Serum | Inquiry |


