Escherichia coli (ETEC) stands as a major food and waterborne pathogen which causes most cases of traveler's diarrhea and severe diarrhea in children from resource-poor areas. ETEC infections lead to 45% of all cases of traveler's diarrhea which occur worldwide. The pathogenic mechanism of ETEC infection depends on multiple essential virulence factors. ETEC uses multiple colonization factors and adhesins to establish an initial connection with human intestinal epithelial cells located in the distal small intestine. The release of heat-labile (LT) enterotoxins and heat-stable (ST) enterotoxins leads to severe watery diarrhea and dehydration in patients. The ETEC infection leads to immediate diarrheal symptoms and creates long-term gastrointestinal problems that result in functional disorders and irritable bowel syndrome.
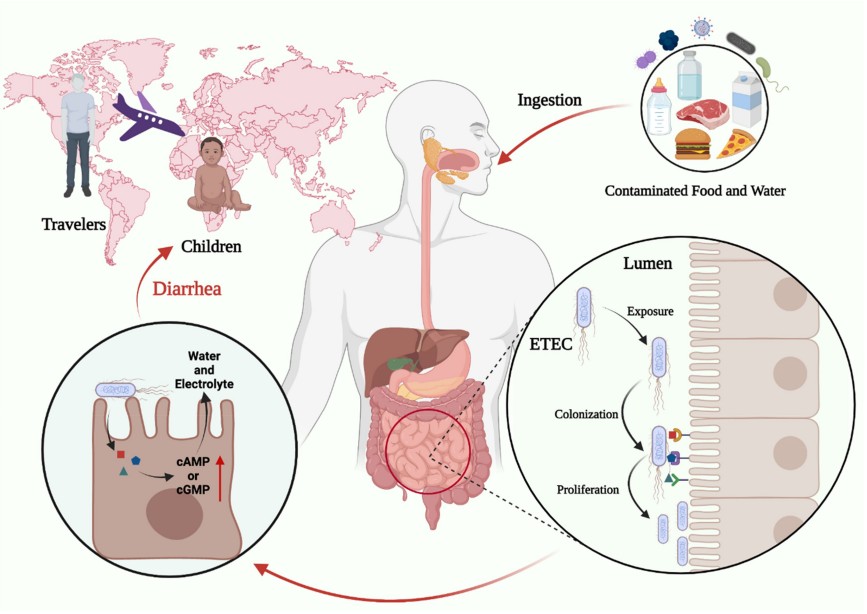 Figure 1. Characteristics of the ETEC infection. (Zhang, 2022)
Figure 1. Characteristics of the ETEC infection. (Zhang, 2022)
ETEC functions as the primary cause of traveler's diarrhea and childhood diarrhea in developing nations and tropical regions including Latin America and the Caribbean and Southern Asia and Africa. The main symptoms of ETEC infection include watery diarrhea together with abdominal pain and nausea and vomiting and fever. The symptoms will last about 3–5 days. ETEC infection is acquired by ingestion of contaminated food or water. Due to the high infectious dose, ETEC are spread by contaminated food and water, not by human-to-human transmission. Oral rehydration therapy and antibiotic therapy are very effective for ETEC infection diarrhea. In recent years, many reports have indicated that antimicrobial resistant ETEC strains are increasing worldwide. ETEC pathogens represent a major health concern for both humans and farm animals. There are at least 78 detectable O antigens and 34 H antigens associated with ETEC. The pathogen is transmitted between humans and/or animals through the fecal-oral route, by ingestion of contaminated food and water exposed to animal and/or human sewage.
Serological typing of ETEC strains have relied on the composition of outer membrane proteins and, mainly, in the somatic LPS (O) and flagellar (H) antigens. Serological typing of ETEC strains have relied on the composition of outer membrane proteins and, mainly, in the somatic LPS (O) and flagellar (H) antigens. ETEC comprise more than 100 somatic serogroups (O) and at least 34 flagellar types (H), combined in an unpredicted number of O:H serotypes, but only a limited number of serotypes are associated with infectious diseases, such as O8:H9, O6:H16, O78:H12 and O25:H42, and are therefore of major clinical relevance.
The pathogenic mechanism of ETEC begins with its oral ingestion, leading to a sophisticated sequence of events that culminate in the release of diarrheagenic toxins in the small intestine. This process relies on a coordinated interplay of plasmid-encoded virulence factors and chromosomally conserved traits, facilitating the bacterium's adherence, colonization, and toxin delivery at the epithelial surface.
 Figure 2. Key steps of ETEC pathogenesis. (Roussel, 2019)
Figure 2. Key steps of ETEC pathogenesis. (Roussel, 2019)
ETEC starts its infection process after a person consumes more than 107 CFU of bacteria through contaminated food or water sources. The stomach environment with its pH of 2 represents the first stage for ETEC to survive before it reaches the small intestine where it can colonize. The bacteria express acid resistance genes (ARs) to survive the stomach environment which includes the glutamate-dependent AR2 and arginine-dependent AR3 systems found in E. coli. The pH level affects ETEC gene expression which controls infection mechanisms including the production of heat-labile toxin (LT) through the eltB gene. The pancreas produces bicarbonate which neutralizes stomach acid during duodenal transit allowing ETEC to survive better. The intestinal epithelium contains a single mucus layer composed mainly of MUC2 which differs from colon mucus because it allows bacterial penetration. The MUC glycoproteins create a hydrated gel-like barrier which protects the intestinal epithelium but ETEC uses mucinases to break down mucins and decrease mucus viscosity for reaching epithelial cells. The essential part of ETEC infection requires the pathogen to bind specifically to intestinal epithelial cells through CFs which function as adherence factors. The pathogen uses these factors to create close contact with host cells which enables successful colonization and toxin delivery. The pathogen uses plasmid-encoded fimbrial and non-fimbrial and fibrillar structures to bind with enterocyte receptors for establishing effective colonization. The adhesins EtpA and EaeH together with other adhesins help ETEC establish successful colonization.
 Figure 3. Schematic representation of the infection cycle of ETEC in the human host. (Salvador-Erro, 2025)
Figure 3. Schematic representation of the infection cycle of ETEC in the human host. (Salvador-Erro, 2025)
The B subunit of heat-labile toxin (LT) recognizes GM1 gangliosides which exist on the apical surface of enterocytes to initiate binding. The toxin enters the cell and the LTA subunit separates from the rest of the toxin to become active in the cytoplasm. The Gsα protein becomes activated through ADP-ribosylation by LTA which then activates adenylate cyclase (AC). The elevated cAMP levels activate protein kinase A (PKA) which phosphorylates the cystic fibrosis transmembrane conductance regulator (CFTR). The increased chloride (Cl−) secretion into the intestinal lumen creates an osmotic imbalance that results in watery diarrhea.
The heat-stable toxin (ST) targets the guanylate cyclase C receptor (GC-C) which exists on the brush border surface of enterocytes. The activation of GC-C results in GTP hydrolysis that produces cyclic GMP (cGMP). The activation of cGMP-dependent protein kinase II (PKGII) by elevated cGMP levels results in CFTR phosphorylation which increases Cl− secretion. The activation of phosphodiesterase 3 (PDE3) by cGMP results in increased cAMP levels which enhance Cl− secretion. The imbalance of ions between the intestinal cells and lumen leads to fluid movement into the intestinal space which produces diarrhea.
 Figure 4. Pathogenesis of ETEC infection. (Joffré, 2015)
Figure 4. Pathogenesis of ETEC infection. (Joffré, 2015)
ETEC strains are characterized by the production of colonization factors (CFs) and at least one of two enterotoxins: LT and ST. Understanding these virulence factors is essential for developing effective vaccine strategies.
Table 1. Virulence factors and involved genes of ETEC.
| Stage of the Infection Cycle | Virulence Factor or System | Genes Involved |
| Survival in the Acidic Environment | AR1 | Unknown |
| AR2 (GDAR) | gadABC, gadEWX, ybaST | |
| Penetration of the Mucus Layer | YghJ (SsIE) | yghJ |
| EatA | eatA | |
| SepA | sepA | |
| Adherence to Intestinal Epithelium | Colonization factors (CFs) | cfaA, cfaB, cs1, cs2, cs3 |
| EtpA | etpA | |
| EaeH | eaeH | |
| Toxin Secretion | LT (heat-labile toxin) | eltA, eltB |
| ST (heat-stable toxin) | estA, estB | |
| EAST-1 (enteroaggregative heat-stable enterotoxin) | astA | |
| HlyA (hemolysin A) | hlyCABD |
The diagnosis of ETEC proves difficult because its symptoms match those of multiple other diarrheal pathogens. The precise identification of ETEC through DNA-based assays remains restricted to limited areas because these advanced diagnostic tools are not accessible in numerous underdeveloped or distant locations. Medical practitioners in these areas must make their diagnoses based on patient histories and symptoms because these methods fail to identify ETEC from Vibrio cholerae and other enteric pathogens. Medical facilities in industrialized nations use stool culture tests along with enterotoxin detection to confirm suspected ETEC infections.
Laboratory methods include: The Polymerase Chain Reaction (PCR) technique serves as a precise method to identify LT and ST toxin genes in samples. The commercial LT toxin detection kits exist for clinical use although they remain uncommon in medical settings. The Enzyme-Linked Immunosorbent Assay (ELISA) used to detect toxins in the past now functions only in research settings because it no longer serves clinical diagnosis needs.
View Heat-labile Enterotoxins (LT) and Heat-stable Enterotoxins (ST)
Pathogenic E. coli: Types, Toxins, and Detection Methods
Enteropathogenic E. coli (EPEC)
Enterotoxigenic E. coli (ETEC)
Enterohemorrhagic E. coli (EHEC)
Adherent invasive E. coli (AIEC)
Enteroaggregative E. coli (EAEC)
Diffusely-adhering E. coli (DAEC)
Verocytotoxigenic / Shiga toxin-producing E. coli (VTEC / STEC)
References
| Target | Cat. No. | Product Name | Host | |
| E. coli | DAG-ZL0485 | Inactivated Escherichia coli ETEC O126:K71 Culture Fluid | N/A | Inquiry |
| DAG-ZL0486 | Inactivated Escherichia coli ETEC O25:K19 Culture Fluid | N/A | Inquiry |
| Target | Cat. No. | Product Name | Host | Isotype | Application | |
| GUCY2C | DMAB-CDB26037 | Hi-Puri™ Mouse Anti-Human GUCY2C Monoclonal antibody, clone PF-07062119 | Mouse | IgG | ELISA | Inquiry |
| DMAB-CDB26038 | Hi-Puri™ Human Anti-Human GUCY2C Monoclonal antibody, clone PF-07062119 | Human | IgG | ELISA | Inquiry | |
| DMAB-CDB26039 | Hi-Puri™ Rabbit Anti-Human GUCY2C Monoclonal antibody, clone PF-07062119 | Rabbit | IgG | ELISA | Inquiry | |
| CAB-3498MH | Anti-GUCY2C monoclonal antibody, clone MAAG945 | Mouse | IgG2b | ELISA, WB | Inquiry | |
| DPAB-DC1512 | Anti-GUCY2C (aa 24-133) polyclonal antibody | Mouse | WB, ELISA | Inquiry | ||
| DPABH-20707 | Anti-GUCY2C (aa 982-1067) polyclonal antibody | Rabbit | IgG | IHC-P | Inquiry | |
| DMABZ016 | Anti-GUCY2C monoclonal antibody, clone 2C22 | Mouse | IgG2b, &kappa | ELISA, WB | Inquiry | |
| DMABZ017 | Anti-GUCY2C monoclonal antibody, clone 3G4 [FITC] | Mouse | IgG2b, &kappa | ELISA, WB | Inquiry | |
| DMABZ018 | Anti-GUCY2C monoclonal antibody, clone 3H8 | Mouse | IgG2b, &kappa | WB, ELISA | Inquiry |
| Target | Cat. No. | Product Name | Host | |
| E. coli Heat Labile Enterotoxin | DAG3923 | Recombinant E. coli Heat-Labile Enterotoxin B subunit | Pichia pastoris | Inquiry |