Human cytomegalovirus (HCMV) is a double-stranded DNA virus that belongs to the beta subfamily of herpesviruses and is the DNA virus with the largest genome among human herpesviruses. The full length of the HCMV genome is approximately 240 kb, consisting of unique long (UL) and unique short (US) sequences, with inverted repeats at both ends. HCMV has a complex proteome, and it is estimated that HCMV has more than 200 open reading frames encoding proteins. HCMV replication is tightly regulated by multiple steps, and gene expression can be divided into immediate early, early and late stages. The glycoprotein (GP) rich in the HCMV envelope is the main determinant of its tissue affinity and plays an important role in the virus entering cells and spreading between cells.
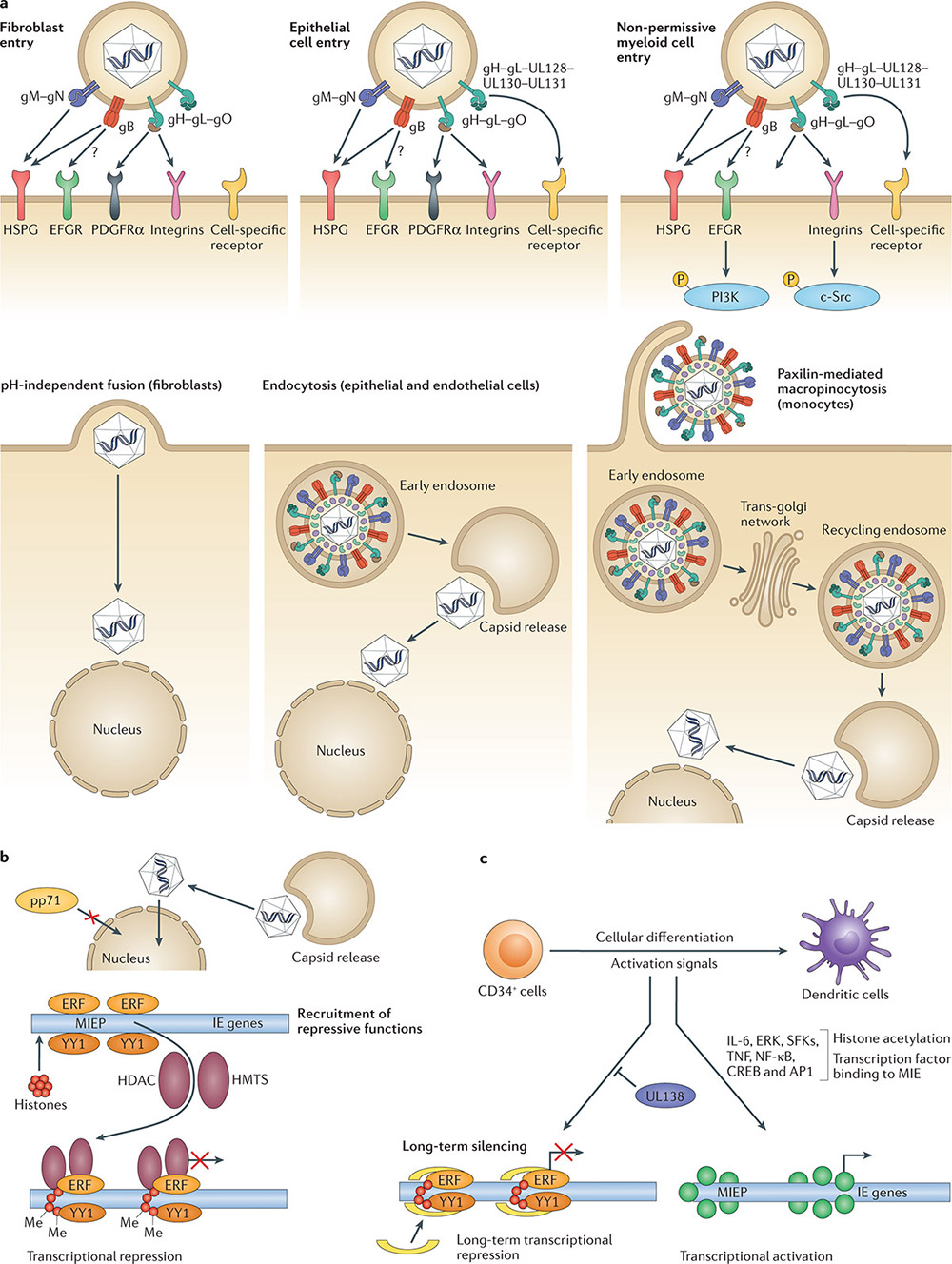 Figure 1. Human cytomegalovirus entry into target cells and the establishment of latency in non-permissive myeloid cells.(Griffiths P, et al.; 2021)
Figure 1. Human cytomegalovirus entry into target cells and the establishment of latency in non-permissive myeloid cells.(Griffiths P, et al.; 2021)
After the body is first infected with HCMV, the virus cannot be completely cleared by the body's immune response and enters a latent infection period, where the virus is excreted for a long time or intermittently. HCMV has various latent sites, and the viral DNA has been detected in monocytes, dendritic cells, megakaryocytes, myeloid progenitor cells and endothelial cells in the bone marrow. In immunocompromised people (including organ transplant recipients, hematopoietic stem cell transplant recipients, HIV-infected patients, and patients receiving immunomodulatory drugs), latent HCMV infection is more likely to be reactivated and invade the lungs, gastrointestinal tract, and central nervous system, and retina and other organs and tissues, causing serious complications and even death. Cytomegalovirus (CMV) infection can be transmitted through multiple ways such as close contact, blood or tissue exposure, perinatal exposure, and sexual exposure. The innate immune barrier is the body's first line of defense against CMV invasion. When the barrier is damaged or if the development is not perfect (such as blood-fetal barrier), the body will be damaged by CMV. During the infection process, CMV is first recognized by non-specific immune cells such as natural killer (NK) cells, monocyte-macrophages, and dendritic cells (DC), initiating the innate immune response. Innate immunity can recognize pathogens, trigger cell signaling cascades, regulate the body's production of interferon (IFN) and pro-inflammatory factors, thereby triggering the body's antiviral response and activating the adaptive immune response. The adaptive immune response can generate protective immunity against CMV through the direct antiviral effect of specific lymphocytes and the secretion of various cytokines, and control virus replication.
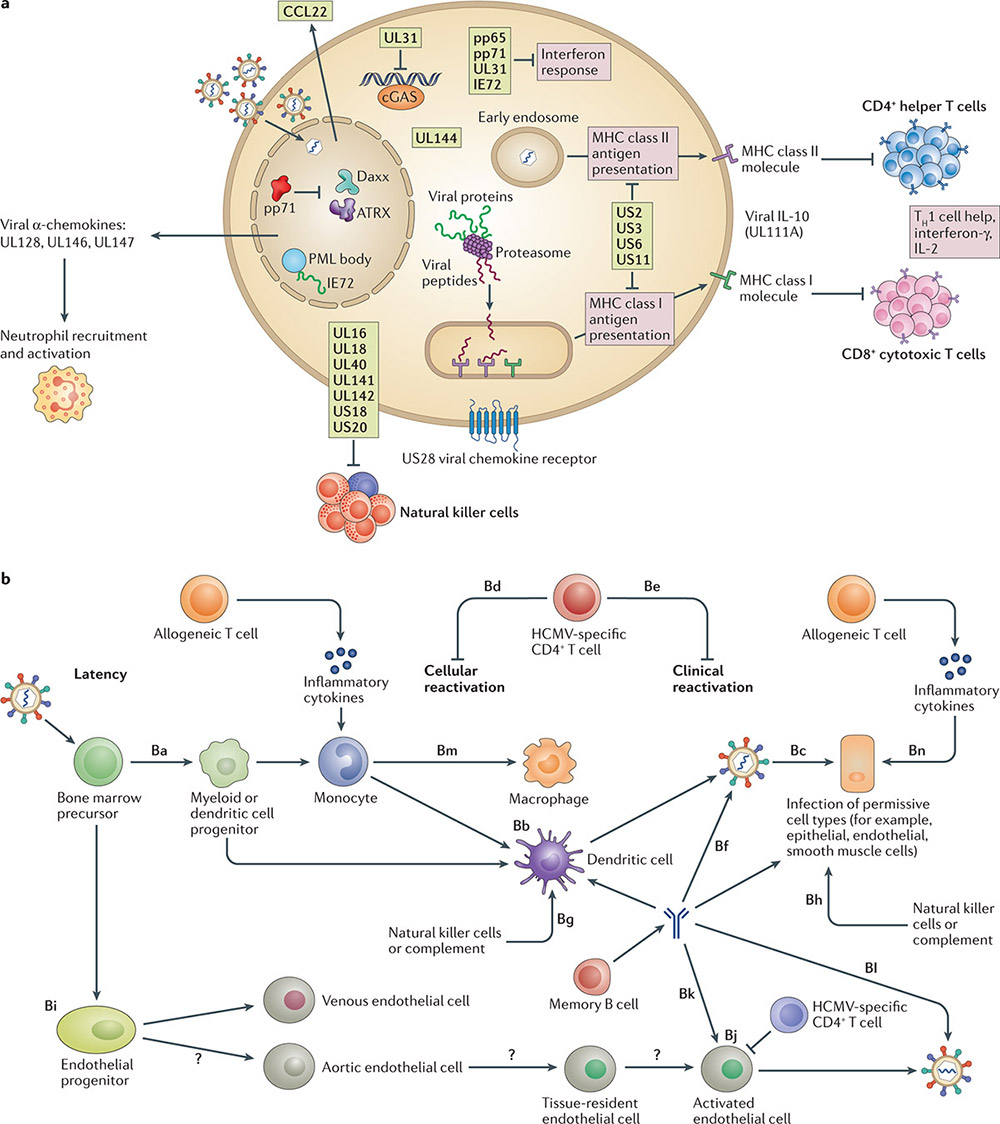 Figure 2. Viral and host functions in human cytomegalovirus latency and reactivation. (Griffiths P, et al.; 2021)
Figure 2. Viral and host functions in human cytomegalovirus latency and reactivation. (Griffiths P, et al.; 2021)
CMV has formed a complex anti-immune response mechanism during its long-term coexistence with the host. It can not only use its huge genome to induce the synthesis of multiple components to participate in the anti-infection response of the host, but also regulate host cells to achieve immune evasion. The immune escape mechanism is an important research topic in the field of CMV. Analyzing its mechanism is of great scientific significance for the design of safe and efficient new vaccines and the development of antiviral drugs. Now we will introduce the research on CMV immune evasion mechanism.
Interference The major histocompatibility complex (MHC) is a major tissue antigen system that represents individual specificity and participates in the regulation of immune responses. It can be divided into three categories according to the structure and function of the encoded protein: MHC-I, MHC class II molecules and MHC class III molecules. Human MHC is also called human leukocyte antigen (HLA). HLA-Ⅰclass molecule is a glycoprotein composed of non-covalently linked α chain (heavy chain) and β chain (light chain). The α chain is encoded by HLA-A, B or C allele, and the β chain is encoded by β2 microglobulin gene encodes, HLA-Ⅰmainly presents endogenous peptides in cells, recognized by CD8+ T cells, and expressed on the surface of all cells in the body except red blood cells; HLA-Ⅱclass molecules are also composed of α chain and β chain Heterodimeric glycoproteins, linked by non-covalent bonds, are encoded by human leukocyte antigen-DR (HLA-DR), DP and DQ genes. They mainly present foreign peptides and are recognized by CD4+ T cells, only expressed on the surface of antigen-presenting cells such as macrophages, DC cells and B cells.
Due to long-term exposure to persistent antigen stimulation and inflammation in chronic infections, T cells gradually lose their effector functions and memory T cell characteristics also begin to be lost, which is called T cell exhaustion. It is mainly manifested by the gradual loss of effector functions (such as defective cytokine production), increased and sustained expression of multiple inhibitory receptors, metabolic disorders, and changes in cellular transcription and epigenetics. The increase in soluble immunosuppressive factors and defective T cell function are the main causes of T cell exhaustion.
IFN can be divided into 3 types: type I, type II and type III IFN. Among them, type I IFN, such as IFN-α and IFN-β, can be secreted by most cells; type II IFN only contains one member, IFN-γ, and is mainly secreted by natural killer (NK) cells and T lymphocytes; type III IFN It is composed of IFN-λ1, IFN-λ2, IFN-λ3 and IFN-λ4. Pattern recognition receptors such as Toll-like receptors, cyclic GMP-AMP synthase (cGAS), and stimulator of interferon gene (STING) can recognize pathogen-related molecular patterns and initiate cell signaling cascades response, promoting IFN gene activation, and subsequent expression and secretion, thus exerting antiviral effects. Interestingly, some studies found that CMV-specific T cell immune testing was performed on severely ill non-immunosuppressed patients, and the results showed that IFN-γ was inversely proportional to CMV DNA load. In addition, the researchers found that lower levels of IFN-γ were associated with a higher risk of developing active CMV infection. Therefore, IFN may be a key factor in inhibiting CMV reactivation.
CMV can evade the killing effect of NK cells by regulating the inhibitory or activating signals of NK cells. Inhibitory receptors include CD94/NKG2C, immunoglobulin-like transcript receptors, etc. among killer cell lectin-like receptors, while NKG2D and NKp30 and others are activating receptors. Multiple gene loci in the CMV genome, such as UL135, UL142 and UL148, have been proven to inhibit NK cell recognition and killing through different mechanisms. UL135 inhibits the formation of immune synapses by remodeling the actin cytoskeleton; glycoproteins encoded by UL142 and UL148A inhibit NK cell activation by downregulating the expression of the NKG2D ligand MICA on the cell surface; UL148 affects the CD2 costimulatory molecule CD58 (LFA-3) expression. In addition, the m02 and m145 family members in CMV can also express MHC-class I molecular analogues, which bind to NK cell inhibitory receptors, inhibit NK cell activation, and protect infected cells from being killed.
DC is the main antigen-presenting cell. Its membrane surface is rich in MHC-Ⅰ, MHC-Ⅱ, co-stimulatory molecules (CD80, CD86 and CD40, etc.), intercellular adhesion molecules and lymphocyte function-related molecules, etc., stimulating initial T cell activation. Proliferation plays an important role in both initial and sustained immune responses. At present, DCs that exist in lymphoid tissues, blood and other non-lymphoid tissues are collectively called classical DCs (conventional DCs, cDCs); DCs that can secrete large amounts of IFN-I are called plasmacytoid DCs (pDCs), which have Different phenotypes and functions. Studies have found that HCMV infection inhibits the maturation of DCs, and after mice are infected with CMV, the fluid-phase endocytosis ability of DCs is impaired and they lose the ability to capture antigens. Studies have confirmed that DCs play a dual role during CMV infection in mice. While generating immune responses, they specifically downregulate antigen presentation to initial T cell surface proteins and initiate immune suppression.
CMV can directly interfere with viral antibody production and effector functions. Studies have found that the glycoproteins gp34 and gp68 encoded by the CMV RL11 gene and UL118-UL119 gene can compete with FcγR on immune cells to bind to the IgGC end, blocking the viral protein-IgG complex from binding to receptors on the surface of phagocytes and NK cells. These processes block antibody-dependent cell-mediated cytotoxicity and the classic complement activation pathway. Interestingly, studies have found that deletion of these two genes significantly enhances the ability of anti-CMV polyclonal IgG antibody-mediated FcγR activation, the ability of immune cells to clear viruses is enhanced.
Reference
| Target | Cat. No. | Product Name | Size | Species Reactivity | Application | Detection Sample | |
| CMV | DEIA-S21CM | CMV IgM ELISA Kit | 96T | Human | Qualitative | serum | Inquiry |
| DEIA013 | Cytomegalovirus IgG ELISA Kit | 96T | Human | Qualitative | serum | Inquiry | |
| DEIA326 | Cytomegalovirus IgG ELISA Kit | 96T | Quantitative | serum, plasma | Inquiry | ||
| DEIA462 | Human CMV IgA ELISA Kit | 96T | Human | Quantitative | Serum | Inquiry | |
| DEIA463 | Human CMV IgG ELISA Kit | 96T | Human | Semi-quantitative | immunocompromised patients | Inquiry | |
| DEIA464 | Human CMV IgG(CSF) ELISA Kit | 96T | Human | Semi-quantitative | Serum | Inquiry | |
| DEIA466 | Cytomegalovirus (CMV) IgM ELISA | 96T | Human | Semi-quantitative | Human serum or EDTA, heparin or citrate plasma | Inquiry | |
| DEIA401 | CMV IgG ELISA Kit | 96T | Human | Semi-quantitative, Qualitative | serum, plasma | Inquiry | |
| DEIA402 | Human CMV-IgM(cytomegalovirus-Immunoglobulin M) ELISA Kit | 96T | Human | Qualitative | Serum, plasma | Inquiry | |
| DEIA2274 | Human CMV IgM ELISA Kit | 96T | Human | Qualitative | serum | Inquiry | |
| DEIA2275 | Human CMV IgG ELISA Kit | 96T | Human | Quantitative | Serum | Inquiry | |
| DEIA465 | Human CMV IgG and IgG avidity ELISA kit | 96T | Human | Qualitative | Serum, plasma | Inquiry | |
| DEIA326R | Human CMV IgG ELISA RUO | 96T | Human | Quantitative and qualitative | serum and plasma (EDTA, lithium heparin or citrate plasma) | Inquiry | |
| DEIA2275S | Cytomegalovirus IgG ELISA kit | 96T | Human | Quantitative and Qualitative | Serum, Plasma and Cell Culture Supernatants | Inquiry | |
| DEIA-NS2307-106 | Human CMV-IgG(cytomegalovirus-Immunoglobulin G) ELISA Kit | 96T | Human | Quantitative | serum, plasma, tissue homogenates and other biological fluids. | Inquiry | |
| DST-H018 | (CMV) Cytomegalovirus IgM/IgG Antibody Rapid Test | 96T | Human | Qualitative | Serum, plasma | Inquiry | |
| DEIAPV4 | Cucumber Mosaic Virus (CMV) ELISA Kit | 500T/ 1000T/ 5000T | Qualitative | host plans | Inquiry |