Epstein-Barr virus (EBV), the most ubiquitous human virus, was first identified in Burkitt's lymphoma (BL) in 1964 and was later identified as the first human oncolytic virus. It has been implicated in the pathogenesis of Hodgkin's lymphoma (HL), non-lymphoid neoplasms in post-transplant patients and HIV-infected individuals, malignancies such as nasopharyngeal and gastric cancers, and in adolescents EBV is the leading cause of infectious mononucleosis (IM). EBV is also known as Human Herpesvirus 4 (HHV-4), and its life cycle is similar to other herpesviruses, with incubation and cleavage periods. Viruses in the latent phase exist as circular segments in the nucleus, linked to host genomic chromatin by the EBNA1 viral protein, and viruses in this state express a small fraction of viral genes and noncoding RNAs. During the latent phase, the virus relies on normal cell division processes, including the host DNA replication machinery, to passively replicate its viral genome and spread to daughter cells. EBV is capable of lifelong latency in the human body, and under certain conditions, the virus is able to reactivate and enter a cleavage cycle in which all viral genes are expressed, and the virus can replicate its viral genome and produce infectious offspring, a process that leads to cell death and the release of infectious viruses, which can infect naïve cells in the same host, or spread to new hosts.
EBV can infect a variety of cells and tissues, including T lymphocytes and B lymphocytes, nasopharyngeal and oropharyngeal squamous cells, salivary and gastric glands, thyroid gland epithelial cells, smooth muscle and follicular dendritic cells. EBV mainly infects and replicates in the layered squamous epithelium of the oropharynx and establishes latent infection in B lymphocytes. In normal carriers, the virus persists in circulating memory B cells and begins to produce immunoglobulins. EBV infection of B cells expresses specific latency-associated genes and transcripts, and the virus remains dormant in quiescent memory B cells and reactivates intermittently at any mucosal site where B cells are present. Researchers estimate that about 1 to 50 out of every million circulating B cells in a healthy adult are infected with EBV. Thus, the EBV virus coexists with most human hosts.
EBV infection of B lymphocytes leads to two outcomes. The first results in the production of long-lasting memory B cells, and the second results in the differentiation of B cells into programmed-death plasma cells. Latently infected B cells express different types of RNAs and proteins, such as EBV-encoded small RNA 1 (EBER1) and EBER2, EBV stabilizing intron sequence RNAs (EBV-sisRNAs), EBV nuclear antigen 1 (EBNA1), and latent membrane protein 1 (LMP1). The type, differentiation, and activation status of infected B cells define different expression programs for EBV-encoded proteins. During the latent phase, viral proteins are reduced to evade recognition of infected cells by cytotoxic T cells. In the second outcome, the virus turns on lysis for replication and simultaneously expresses multiple viral proteins, including the trans-activating protein BZLF1 (also known as ZEBRA) and the viral protein complexes (Early Antigen (EA) and Viral Capsid Antigen (VCA)), thereby eliciting a humoral immune response. Regulatory proteins are sensitized thereby producing viral DNA (EBV-DNA), VCA and membrane proteins (MAs).
Table 1. EBV in infected B-cells with EBV latency pattern and associated malignancy
| Infected Cells | |||||
| Native B-cells | Germinal Center B-cells | Peripheral Memory B-cells | Dividing Peripheral Memory B-cells | Plasma Cells | |
| Transcription program | Latency Ⅲ | Latency Ⅱ | Latency 0 | Latency Ⅰ | Lytic |
| Viral proteins | All EBNAs, EBERs, LMP-1, LMP-2A and LMP-2B | EBNA-1, EBERs, LMP-1 and LMP-2A | EBERs | EBNA-1 and EBERs | All lytic genes |
| Function of viral proteins | Activate B-cell | Differentiate activated B-cell into memory B-cell | Allow for lifetime persistence | Allow for the virus in latency-programmed cell to divide | Assist viral replication in plasma cells |
| Associated malignancies | IM and post-transplant lymphoproliferative disorder | Nasal NK cell lymphoma, Hodgkin's lymphoma, chronic active EBV infection, NPC and peripheral NK/T cell lymphoma | Healthy carrier | Burkitt lymphoma and gastric carcinoma | IM and NPC |
| Specimens for measuring viral load | Plasma or serum, MNCs and WBC | Plasma or serum, MNCs (for chronic active EBV infection), tissue biopsy | Plasma or serum, WBC | Plasma or serum | Plasma or serum |
EBV, Epstein–Barr virus; EBNA, Epstein–Barr virus nuclear antigen; LMP, latent membrane protein; EBERs, EBV-encoded small RNAs; NK cells, natural killer cells; NK/T cell, nasal natural killer (NK)/T-cell; MNCs, mononuclear cells; WBC, white blood cell; IM, infectious mononucleosis; NPC, nasopharyngeal carcinoma.
(Source: AbuSalah MAH, et al. 2020)
The lysis form of EBV infection is required to produce infectious viral particles and is essential for cell-to-cell and host-to-host transmission of the virus. The number of EBV-infected B cells decreases after the use of drugs to inhibit lysis of EBV replication, suggesting that reactivation and replication of EBV may help to replenish the pool of latently infected cells in the infected host. Lytic EBV genes are categorized as IE, early or late lytic genes, and lytic reactivation is mediated by the expression of two viral IE lytic proteins, BZLF1 (Z, ZTA, ZEBRA) and BRLF1 (R, RTA). Z and R encode viral transcription factors that collectively activate each of the early lysis EBV proteins required for replicative lytic forms of EBV viral DNA expression. The early cleavage gene promoter contains cis-acting motifs that bind to Z and/or R. Transcription is activated when bound to Z and/or R proteins. Late cleavage genes encode proteins required for viral particle assembly. The activity of the viral promoters that drive Z and R expression (Zp and Rp, respectively) is mainly regulated by cellular transcription factors. Important biological inducers of B-cell lytic EBV reactivation include B-cell receptor activation and plasma cell differentiation. In addition, remaining stimuli, such as hypoxia, inflammasome induction and DNA damage, can also induce EBV reactivation. In contrast, other cellular transcription factors (ZEB1/2, c-myc, KAP1, and IRF4) promote viral latency.
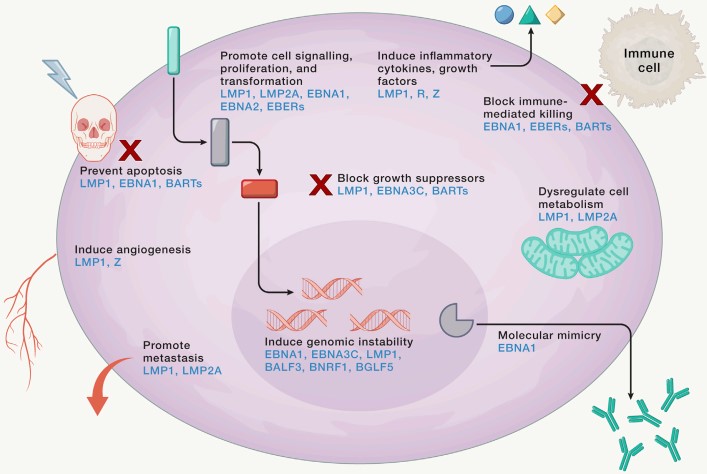 Figure 1. EBV modulation of the infected cell
Figure 1. EBV modulation of the infected cell
(Source: Damania B, et al. 2022)
Prior to the discovery of EBV, chronic active EBV (CAEBV) infection was described as chronic IM, and severe CAEBV was considered a well-defined disease entity because of its characteristic clinical, virologic, and pathologic findings and extremely poor prognosis. Abnormally elevated IgG antibodies against EBV viral lysate antigens and increased EBV viral replication in patients. In addition, homogeneous and phenotypic EBV populations are often present in affected lesions or peripheral blood, suggesting initial EBV infection of single progenitor cells followed by clonal proliferation. Abnormal cellular regulation and immunosurveillance defects or imbalances are also present in patients with CAEBV. Normally the EBV receptor CD21 is expressed on human B-cells and not on T-cells and NK-cells, but there still exists a subset of CAEBV patients who develop EBV genome-positive T-cell lymphomas, a phenomenon that exists only in CAEBV patients.
Primary infection with EBV typically occurs in childhood, with no obvious clinical signs of infection, and when uninfected adolescents are exposed to EBV, approximately 30%-70% will develop IM. For primary infectious mononucleosis, the usual incubation period is 32-49 days, when there are no obvious symptoms. In the acute phase, patients present with a sore throat, headache, cervical lymphadenopathy, fever, fatigue, and in a few cases hepatomegaly, nausea, vomiting, eyelid edema, and rash. A feature of the clinical presentation of IM is the long duration of the disease, with a median acute episode duration of 18 days. Serious complications are rare in the acute phase of primary EBV infection; very few patients may develop airway obstruction due to oropharyngeal inflammation, meningoencephalitis, streptococcal pharyngitis, hemolytic anemia, and thrombocytopenia. When an acute infection moves to the chronic phase, chronic IM can be categorized into two clinical presentations, the first of which involves a gradual return to health, but with persistent recurrent symptoms months to years later. The second pattern is rarer and presents as a persistent "monotypic" disease that continues indefinitely.
From a clinical point of view, EBV testing will help to make a correct diagnosis for the patient, contribute to therapeutic monitoring and prognosis of EBV-associated diseases, and in the diagnosis of EBV-associated malignancies EBV can also be used as a biomarker for the diagnosis and assessment of tumor spread as well as for monitoring therapy. Laboratory tests for the accurate detection of EBV are also important from the point of view of basic and epidemiologic research. Several methods are currently used for EBV testing, including serologic and molecular diagnostic methods, each of which has its advantages and limitations. In situ hybridization (ISH) is the gold standard method for detecting EBV-associated cancers, but molecular assays of viral DNA, RNA, and EBV viral load are now being used in the clinical evaluation of tumor-associated EBV infection. Point-of-care testing is usually based on the detection of heterophilic antibodies, which are produced during the systemic immunoregulatory process that accompanies acute primary EBV infection and are IgM-like antibodies directed against mammalian erythrocytes. The most useful EBV-specific antibody tests are VCA IgM, VCA IgG, and EBNA-1 IgG, which are usually detected by immunoassay platforms. Antibodies against EBNA-1 are generally used in commercial diagnostic reagents, but the antibodies are slow to be produced and are usually not detectable until 3 months or more after onset of disease.
Table 2. Advantages and disadvantages of various EBV diagnostic methods
| Method | Advantages | Disadvantages |
| Molecular methods (PCR and other nucleic amplification methods) | (1) Ability to differentiate between healthy carriers and patients with EBV-related disease based on viral load (low or high) (2) Low risk of contamination and reduced labor costs and turnaround time in qPCR (3) Allow for quantitative EBV DNA detection to monitor disease status. (4) For early intervention, it is useful in screening high-risk populations and in monitoring EBV reactivation (5) Sensitive and specific across a wide dynamic range | (1) Could generate false-positive results due to improper blood sample storage and false-negative results due to the presence of nucleases (2) Lack of standardization (3) Expensive (4) Require special equipment |
| ISH | (1) Ability to identify EBV DNA or EBER transcripts in EBV–associated tumors. (2) Highly reliable confirmatory test for EBV (gold standard for EBV diagnosis) | (1) Only applicable to cells (2) Requires special skills (3) Could get counterproductive due to the histological interference between non-Hodgkin's and Hodgkin's lymphoma (4) EBER is downregulated in oral hairy leukoplakia |
| Heterophile antibody test | (1) Can measure heterophile antibodies released against serum viral proteins (2) Can differentiate between late primary infection and reactivation (3) Cost effective and easy to perform | (1) Less sensitive and less specific (especially in children) (2) Possibility of false-positive result in some cases of autoimmune disease (3) Possibility of false negative is high in young children |
| IFA (immunofluorescence assay) | (1) Gold standard reference method (2) Highly specific (3) Allows for the staging of EBV infections | (1) A high degree of variability (2) Lacks standardization (3) Equivocal diagnosis of acute EBV infection |
| EIAs and ELISA | (1) Rapid method (2) More sensitive than the IFA (3) Suitable for automation (4) Inexpensive (5) Less hands-on time | (1) Less specific (2) Difficulty in the staging of EBV infection (single patient's serum) (3) Lack of standardization (4) Equivocal diagnosis of acute EBV infection |
(Source: AbuSalah MAH, et al. 2020)
EBV is associated with a variety of lymphatic malignancies as well as epithelial cancers such as nasopharyngeal and gastric cancers. EBV is an important factor in these pathologic processes, combining genetic, environmental, immunologic, or infectious factors to lead to cancer development. Different types of cancers are associated with EBV to different degrees; for example, EBV is associated with nasopharyngeal carcinoma, endemic BL, T/NK lymphoma, and HIV-associated primary central nervous system lymphoma in 100% of cases, whereas it is associated with HL, NHL, and gastric cancer in less than 30% of cases. EBV is associated with undifferentiated nasopharyngeal carcinoma (NPC), and some investigators have suggested that persistent infection of genetically aberrant epithelial cells with EBV induces tumorigenic transformation and clonal expansion of infected cells. Other cofactors, such as long-term exposure to environmental carcinogens, contribute to the development of nasopharyngeal carcinoma and can induce a range of somatic genetic changes in epithelial cells. EBV DNA molecules circulating in the blood of NPC patients in the form of naked DNA fragments released from cancer cells into the plasma are considered prognostic biomarkers for nasopharyngeal carcinoma, with potential clinical value in screening, prognosis, and recurrence monitoring.
References
| Target | Cat. No. | Product Name | Size | Species Reactivity | Application | Detection Sample | |
| EBV | DEIABL336 | Epstein Barr Virus (EBV) EA IgM ELISA Kit | 96T | Human | Semiquantitative | Serum or plasma | Inquiry |
| DEIABL337 | EBV VCA IgM µ-capture ELISA Kit | 96T | Quantitative, Qualitative | Serum, plasma | Inquiry | ||
| DEIA1713 | Human EBNA IgG ELISA Kit | 96T | Human | Qualitative | Human sera | Inquiry | |
| DEIA1922 | Epstein Barr Virus(EBNA-1) IgM ELISA Kit | 96T | Human | Quantitative | Serum | Inquiry | |
| DEIA1030 | Epstein Barr Virus NA1 IgA ELISA Kit | 96T | Human | Qualitative | Serum, plasma | Inquiry | |
| DEIA1034 | Epstein Barr Virus Zta IgA ELISA Kit | 96T | Human | Qualitative | Serum, plasma | Inquiry | |
| DEIA3372 | Human EB-VCA-IgG(Epstein-barr virus capsid antigen-Immunoglobulin G) ELISA Kit | 96T | Human | Qualitative | Serum, plasma | Inquiry | |
| EBV VCA | DEIA470 | Human VCA EBV (CSF) IgG ELISA Kit | 96T | Human | Quantitative | Serum | Inquiry |
| DEIA335 | Human EBV VCA IgG ELISA Kit | 96T | Human | Quantitative | Serum, plasma | Inquiry | |
| DEIA336 | Anti-VCA EBV IgA ELISA Kit | 96T | Human | Quantitative | Serum | Inquiry | |
| DEIA337 | Human EBV VCA IgM ELISA Kit | 96T | Human | Quantitative | Serum, plasma | Inquiry | |
| EBV EA | DEIA-NS2310-9 | Epstein-Barr Virus EA IgG ELISA Kit | 96T | Human | Quantitative | Serum or plasma | Inquiry |
| EBV EBNA1 | DEIA332 | Human EBV EBNA1 IgG ELISA Kit | 96T | Human | Quantitative | Serum, plasma | Inquiry |
| Target | Cat. No. | Product Name | Expression System | Tag/Conjugate | Application | |
| EBV | DAG-WT345 | Recombinant EBV NA1 (a.a. 1-90, 408-498) [His] | E. coli | His | WB/ELISA | Inquiry |
| DAG3069 | Recombinant EBV type 1 (a.a. 340) | P. pastoris | Unconjugated | ELISA | Inquiry | |
| DAG1577 | Recombinant EBV Mosaic EBNA1 Protein [GST] | E. coli | GST | ELISA, WB | Inquiry | |
| DAG1578 | Recombinant EBV [His] | E. coli | His | ELISA, WB | Inquiry | |
| DAG1580 | Recombinant EBV p18 Protein fus [GST] | E. coli | GST | ELISA, WB | Inquiry | |
| EBV Nuclear Antigen | DAG-T1129 | EBV Nuclear Antigen (aa 1 - 119) | E. coli | Unconjugated | N/A | Inquiry |
| DAG-P2288 | EBV Nuclear Antigen (aa 1 - 352) | E. coli | Unconjugated | WB, ELISA, SDS-PAGE | Inquiry | |
| EBV Early Antigen | DAG-P2607 | EBV EA regions (full length) | E. coli | Unconjugated | ELISA, WB | Inquiry |
| EBV VCA | DAG3084 | Native EBV VCA Glycoprotein 125 | N/A | Unconjugated | ELISA, WB | Inquiry |
| DAG4598 | Recombinant EBV VCA p23 | E. coli | His | N/A | Inquiry | |
| EBV BMRF1 | DAG1849 | Recombinant EBV BMRF1 [GST] | E. coli | GST | SDS-PAGE | Inquiry |
| EBV EBNA1 | DAG1850 | Recombinant EBV EBNA1 [GST] | E. coli | GST | SDS-PAGE | Inquiry |
| DAG-T1130 | Recombinant EBV EBNA1 mosaic Protein (a.a. 1-90, 408-498) | E. coli | Unconjugated | N/A | Inquiry |