There are four known types of human T-cell leukemia virus (HTLV): HTLV-1, HTLV-2, HTLV-3 and HTLV-4. HTLV-1 is most pathogenic to humans, while HTLV-2 usually causes mild neurological disease. Both viruses are globally endemic. HTLV-1/2 belongs to the retroviral family and is a spherical enveloped virus with an envelope containing glycoproteins, which are important for the binding of the virus to cellular receptors and for the fusion of the envelope with the cell membrane. The protein capsid contains the viral genome, which contains two RNA molecules, protease, reverse transcriptase (RT), integrase, and RNAse H. These enzymes promote viral replication, and RT is responsible for transcribing single-stranded RNA into double-stranded DNA molecules, which integrate into the genome of the host cell and become proviral DNA.
HTLV-1 belongs to the subfamily of orthoretroviruses and was the first human retrovirus to be discovered, with endemic infections in Africa, South America, the Caribbean, southwestern Japan, and the Pacific islands. Human infection with the virus has the potential to induce adult cell leukemia (ALT), HTLV-1-associated myelopathy/tropical spastic paraplegia (HAM/TSP), and a number of inflammatory diseases (keratitis, dermatitis, and conjunctivitis). HTLV-1 does not capture proto-oncogenes or induce proviral insertion mutations like other retroviruses, it is a trans-activating retrovirus that encodes auxiliary proteins capable of inducing cellular transformation over a longer period. This pathogenic characteristic of HTLV also results in a unique clinical aspect of its mediated diseases, namely, the clinical latency period of HTLV-1-associated disorders can be as long as several decades.
The HTLV-1 genome is an approximately 9-kb positive-sense RNA genome with two copies of single-stranded RNA in the viral particle. Upon entry into the host cell, the ssRNA genome is reverse transcribed to dsDNA and integrated into the host genome. The integrated retroviral genome in the form of dsDNA is referred to as provirus. The Long Terminal Repeat Sequence (LTR) at the 5' and 3' ends of the viral genome consists of a unique 3 (U3), repeat (R), and a unique 5 (U5) region. The LTR also contains important elements necessary for viral transcription, polyadenylation, and integration, as well as standard structural and enzyme genes (gag, pro, pol, and env) essential for viral replication. All HTLV-1 genes except one (Hbz) are encoded by the sense strand of the proviral genome. HTLV-1 encodes accessory genes that contribute to viral biology, the most critical of which is Tax. Tax proteins are essential for transformation and act as transcription factors to enhance viral transcription from the 5' LTR promoter. In contrast, Hbz proteins encoded by antisense genomic chains counteract many of Tax's activities, including transcriptional activation of the 5' LTR promoter.
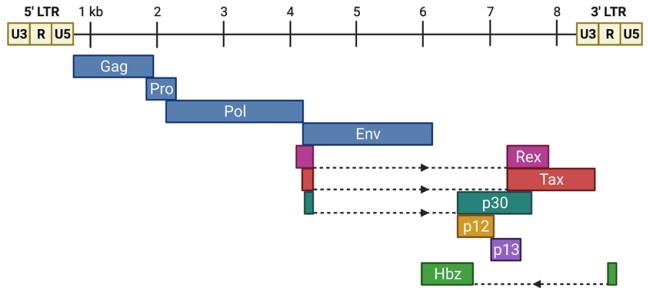 Figure 1. The HTLV-1 proviral genome
Figure 1. The HTLV-1 proviral genome
(Source: Ernzen KJ, et al. 2022)
The Tax protein, expressed by the Tax gene, is an immunodominant antigen with translational and transcriptional activity that is associated with dysregulated immune responses in patients with HTLV-1-associated myelopathy. Tax expression leads to sustained cell proliferation characterized by abnormal expansion of infected cells, producing DNA lesions characteristic of adult T-cell leukemia/lymphoma (ATLL). It also inhibits cellular checkpoint activity to control transcriptional errors, which can lead to the proliferation of DNA-damaged infected cells. Tax is involved in OX40 overexpression, which promotes the proliferation and survival of effector and memory T cells and inhibits the differentiation and activity of T regulatory cells (Tregs). Some studies have found elevated levels of soluble OX40 in the central nervous system (CNS) of patients with HAM, and the expression of Tax in the CNS of patients with HAM leads to the loss of cells capable of activating and generating specific immune responses against TAX, which can directly or indirectly damage neurons. Tax also exerts pleiotropic effects on cell signaling pathways, modulating different cellular pathways and thus primarily promoting HTLV-1-induced tumor transformation.
HTLV-1 is transmitted through breastfeeding, exposure to infected blood products, and sexual behavior. In vivo HTLV-1 relies primarily on cell-to-cell transmission rather than direct infection of target cells by cell-free virosomes, which is partly due to the very limited ability of free HTLV-1 viruses in the blood to infect most cells. Cell-to-cell transmission is common during the acute infection phase of HTLV-1, where the virus is delivered to healthy T cells via infected dendritic cells (DCs), and among the T cell populations, CD4+ T cells are the primary and preferred target cells for HTLV-1 infection, with CD8+ T cells accounting for approximately 5% of the total number of infected cells. The primary pathway by which HTLV-1 establishes persistent chronic infection is through clonal expansion of infected cells after initial infection, rather than de novo infection of cells. Persistent infection with HTLV-1 may be associated with the ability of the virus to evade the host immune response. Immune evasion may be associated with high viral loads and therefore with disease outcome. In addition, HTLV-1 can spread to neighboring cells through the establishment of cellular conduits, formation of virological synapses, and extracellular viral assembly.
 Figure 2. Intercellular transmission of HTLV-1
Figure 2. Intercellular transmission of HTLV-1
(Source: Aghajanian S, et al. 2020)
While HTLV-1 transmitted through breastfeeding or sexual intercourse needs to cross the mucosal barrier to enter the host, virus transmitted through blood products and organ transplantation can be transported directly to the bone marrow and lymph nodes, where physiologic hypoxia increases plus-strand expression, helping to establish a large number of clones before effective antiviral responses can take effect. HTLV-1 can either directly penetrate the damaged epithelium or utilize a cell-mediated pathway. Alternatively, free HTLV-1 viruses can cross the epithelial barrier and infect human dendritic cells under the epithelial barrier via a cytosolic mechanism. As antigen-presenting cells, dendritic cells of different subtypes in the peripheral blood can present antigens to T cells in lymphoid organs, and on the contrary, infected CD4+ T cells can infect dendritic cells through viral synapses, based on the above characteristics, HTLV-1 can establish a bidirectional viral transmission pathway by infecting dendritic cells. Hematopoietic stem cells (HSC) are self-replicating multipotent cells that can differentiate into lymphocytes and myeloid cells in the bone marrow. Identical proviral integration sites between hematopoietic stem cells, leukocytes, platelets, and neutrophils in HAM/TSP suggest that these cells are infected in vivo and produce hematopoietic stem cell-derived HTLV-1 to infect lymphocytes, monocytes, DCs, and other hematopoietic lineages that serve as viral reservoirs in persistent infections.
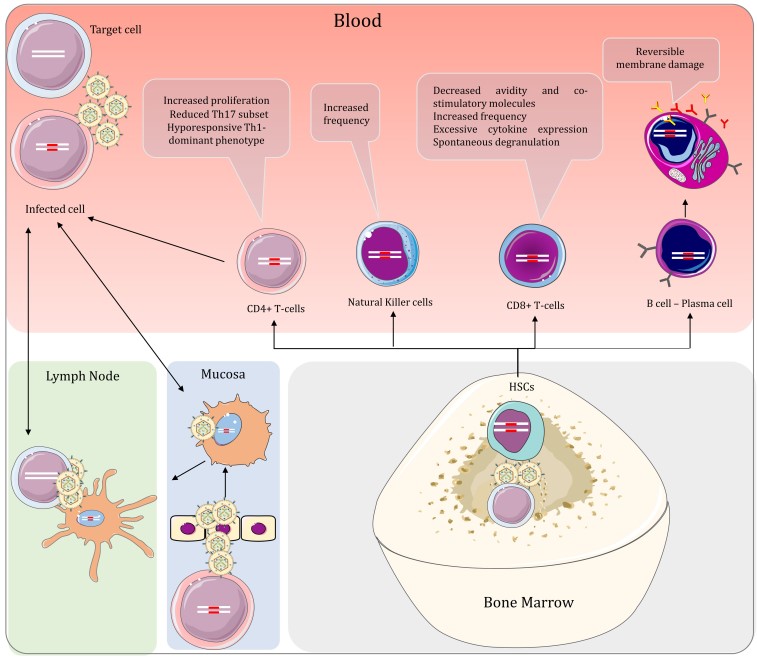 Figure 3. Peripheral interactions of HTLV-1 and the host in HAM/TSP and the effect of the virus on the lymphoid lineage cells
Figure 3. Peripheral interactions of HTLV-1 and the host in HAM/TSP and the effect of the virus on the lymphoid lineage cells
(Source: Aghajanian S, et al. 2020)
Enzyme-linked immunosorbent assay (ELISA) or particle agglutination assay (PA) can initially screen for HTLV-1 infection, and Western Blot (WB) or innogenetics line immunoassay (INNO-LIA) can further confirm HTLV-1 virus. Qualitative or quantitative polymerase chain reaction (PCR) can also be used to detect the HTLV-1 genome, but it is expensive and operator-demanding, and the method is not universally applicable. Samples with recurrent reactions in screening tests must be examined for the presence of antibodies specific to HTLV 1/2 by confirmatory analysis. WB is the reference test for confirmation of infection and determines a positive or negative result for HTLV-1 antibodies. In conclusion, antibody testing is the best choice for routine diagnostic purposes.
HAM/TSP is a chronic hemiplegia with an insidious onset, with lesions occurring primarily in the posterior and lateral columns of the middle and lower thoracic segments of the spinal cord. The disease is characterized by progressive demyelination and neuronal loss in the central nervous system, with characteristic pathology showing spinal cord atrophy and perivascular and parenchymal mononuclear cell infiltration in the CNS. Patients present with lower extremity paralysis, lower back pain, autonomic dysfunction, mild sensory impairment, constipation, and cognitive decline. Virus-host immune interactions play a key role in the pathogenesis of HAM/TSP, where HTLV-1-infected T cells and HTLV-1-resistant CD8+ cytotoxic T lymphocytes (CTLs) release proinflammatory cytokines and chemokines in the CNS, leading to tissue damage. These immune responses are thought to underlie the pathogenesis of HAM/TSP. Tax and HBZ play key roles in the immunomodulatory dysregulation of HAM/TSP. Tax induces the expression of many host cell genes, which promotes cell activation and proliferation. HBZ induces host inflammation by altering the expression of Foxp3 in Treg cells.
The characteristic pathology of HAM/TSP includes chronic inflammation and diffuse degeneration throughout the CNS. The spinal cord shows myelin sheaths and axons bereaved symmetrically in the lateral and posterior columns, with inflammation in both gray and white matter, with a predominance at the level of the thoracic vertebrae. These lesions involve perivascular and parenchymal lymphocytic infiltration with reactive astrocytosis and fibrogliocytosis. Active inflammatory lesions were evenly distributed with CD4+ and CD8+ cells, whereas in the chronic phase CD8+ cells predominated and high levels of IFN-γ were detected. Patients with HAM/TSP have a marked increase in peripherally activated HTLV-1-specific CTL. HTLV-1-specific CTL infiltrate the CNS and play a role in the pathogenesis of HAM/TSP. In addition, large numbers of CD8+ T cells infiltrate the spinal cord and express TIA-1 molecules as CTL markers. The frequency of HTLV-1 Tax-specific CTLs in CD8+ T cells infiltrating the CNS exceeded 20%. In addition, HTLV-1 protein was not detected in CNS-resident cells but was found in more than 60% of infiltrating CD4+ T cells.
References
| Target | Cat. No. | Product Name | Size | Species Reactivity | Application | Detection Sample | |
| HTLV | DEIA079 | Human T-cell lymphotropic Virus antibody, HTLV (1+2) Ab ELISA Kit | 96T | Human | Qualitative | Serum, plasma | Inquiry |
| DEIA10160 | HTLV-I/II p19 Antigen ELISA Kit | 96T | Human | Quantitative | Cell cultures | Inquiry |