The Maedi-Visna virus (MVV) is a small ruminant lentivirus (SRLV) that infects sheep and goats. It is closely related to the caprine arthritis-encephalitis virus (CAEV) and shares genetic, structural, and pathogenic similarities with it. Molecular studies have revealed that these viruses have various variants that can affect both sheep and goats. Interestingly, some variants that typically infect sheep can also infect goats, and vice versa. However, MVV primarily infects sheep, while CAEV is more common in goats, suggesting a preference for infection within the same species. These two types of infection are widespread worldwide.
The clinical manifestations of MVV and CAEV infections can be categorized into four major forms: respiratory, mammary, joint, and neurological. The respiratory form is characterized by interstitial pneumonia, leading to respiratory difficulties in infected animals. The mammary form may result in subclinical mastitis, which can cause a decrease in milk production. The joint form manifests as lameness due to arthritis, affecting the mobility of infected animals. Lastly, the neurological form is characterized by chronic nonpurulent meningoencephalomyelitis, leading to neurological signs such as ataxia and paralysis.
Transmission of MVV and CAEV is primarily through body fluids, specifically respiratory exudates and milk or colostrum, which may contain infected monocytes and macrophages. It is unclear whether transmission occurs through free virions or infected cells. Other modes of transmission, such as transplacental transmission and transmission through semen, are less significant from an epidemiological standpoint. Regardless of the transmission route, the viruses are capable of crossing mucous barriers and infecting macrophages and dendritic cells within the host.
The particles of Maedi-Visna virus (MVV) and caprine arthritis-encephalitis virus (CAEV) have a diameter ranging from 90 to 120 nm. They consist of an envelope and a core. The envelope is composed of a membrane bilayer derived from host cell phospholipids, with viral glycoproteins embedded within it. The core, also known as the capsid, is a protein shell that contains viral RNA and the enzymes required for viral replication. Similar to other retroviruses, MVV and CAEV virions contain two linear RNA molecules. These single-stranded RNA molecules are converted into double-stranded DNA (dsDNA) by the enzyme reverse transcriptase (RT). This mechanism allows the retroviral genome, known as a provirus when it is in the dsDNA form, to integrate into the host cell's genome.
The genomes of MVV and CAEV are approximately 8,400 to 10,000 nucleotides long and consist of three main genes found in all replication-competent retroviruses: gag, pol, and env, along with additional regulatory genes. The proviral DNA is flanked by repeated sequences called long terminal repeats (LTRs), which contain promoter elements responsible for initiating DNA transcription. These LTRs play important roles in cellular tropism, pathogenesis, and gene regulation. The gag gene encodes internal structural proteins, including the capsid (p25CA), matrix (p16MA), and nucleocapsid (p14NC) proteins. The pol gene encodes enzymes involved in replication and DNA integration, such as protease (PR), reverse transcriptase (RT), dUTPase, and integrase (IN). The env gene encodes surface (gp135SU) and transmembrane (gp46TM) glycoproteins inserted into the viral envelope. The accessory regulatory genes vif, vpr-like, and rev are located coinciding with the regions pol and env in different reading frames. These genes contribute to viral replication regulation, infectivity, and evasion of cellular defense mechanisms.
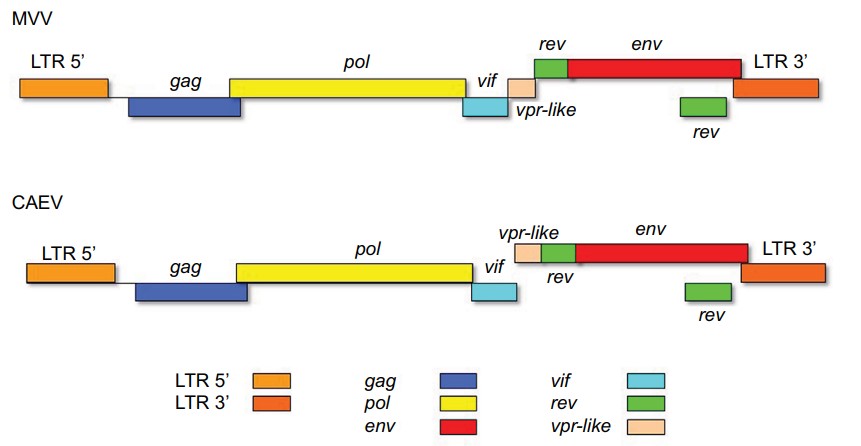 Figure 1. Schematic representation of the gene distribution in the proviral genomes of MVV and CAEV.
Figure 1. Schematic representation of the gene distribution in the proviral genomes of MVV and CAEV.
(Source: Gomez-Lucia, E. et al., 2018)
MVV and CAEV may use different cell receptors for entry into host cells. Recent studies suggest that a mannose receptor serves as the cellular receptor in sheep. Once inside the cell, the replication process is similar to other retroviruses, integrating the viral DNA into the host genome, leading to lifelong infection. The viral envelope is formed from the host cell's lipid membrane, incorporating specific glycoproteins. The host cell deploys innate and adaptive immune mechanisms to combat SRLV infection, such as TRIM5 inhibiting viral integration and APOBEC3 mutating the viral genome. However, retroviruses have strategies to counteract these defense mechanisms.
The U3 region of the LTRs flanking the proviral DNA of SRLVs contains binding sites for cellular factors that activate transcription. These transcription-binding sites (TBS) include ISRE and GAS. Activation of these sites occurs when the cell is stimulated by interferons (IFN), leading to the binding of cellular molecules and modulation of transcription. Similar functional ISRE sites have been found in the 5' LTR of HIV, a closely related lentivirus. Activation of ISRE may play a role in countering host defenses mediated by IFN during the early stages of viral infection.
Reference
| Target | Cat. No. | Product Name | Expression System | Tag/Conjugate | Application | |
| TRIM5 | CDBP6364 | TRIM5 blocking peptide | N/A | Unconjugated | IB | Inquiry |
| CDBP3063 | Human TRIM5 blocking peptide | N/A | Unconjugated | Apuri, BL, ELISA | Inquiry | |
| CDBP3064 | Human TRIM5 blocking peptide | N/A | Unconjugated | Apuri, BL, ELISA | Inquiry |
| Target | Cat. No. | Product Name | Size | Species | Application | Detection Sample | |
| MVV | DEIA457 | MVV/CAEV ELISA Kit | 96T | Sheep, Goat | Qualitative | Serum | Inquiry |
| IFN | DEIA1467 | High Sensitivity Interferon Gamma Human ELISA Kit | 96T | Human | Quantitative | Serum, plasmas, buffered solutions, cell culture media | Inquiry |