Mitochondria are a type of organelle widely present in all types of eukaryotic cells. They were first discovered in animal cells. The shapes of mitochondria are diverse and often vary depending on cell types and physiological conditions. In general, they are Strip or granular shape, "sausage" shape under electron microscope, generally 0.5-1.0 μm in diameter and 2-8 μm in length.
Research results all show that mitochondria are the place where cells perform respiration. Enzymes and coenzymes needed to catalyze the citric acid cycle, amino acid metabolism, fatty acid decomposition, electron transfer, energy conversion, replication and synthesis are all distributed in mitochondria. Their main functions are: 1. Participate in oxidation reactions in the tricarboxylic acid cycle; 2. Electron transfer and energy conversion.
The internal structure of mitochondria has both commonalities and particularities under electron microscope observation. Its commonality is reflected in the fact that each mitochondria has a common basic structure, which can be divided into three parts: the surface of the mitochondria is surrounded by an outer membrane, and there is an inner membrane inside, with many different forms of cristae extending inward, which vary depending on the type, forming a complex The internal system of the cristae is the intracranial space. The electron density of the inner and outer membranes is high. The thickness of each layer is about 5-7nm. The distance between the two is 8.5nm thick. It is an electron-transparent layer called the peripheral space. It is connected with the cristae; 3. The space around these internal systems is filled with a matrix with a smaller electron density, which is a solution containing soluble proteins and other substances. Here we mainly discuss the structure and function of the inner membrane.
The intima is about 6-8nm thick, has a high content of cardiolipin, a low phospholipid to protein ratio (0.27), and a high density (1.2).
In eukaryotic cells, oxidative phosphorylation is mainly performed on the inner mitochondrial membrane. Therefore, the components directly related to energy conversion on the inner membrane can be divided into two categories: 1, belongs to the ATPase Complex (energy conversion device); 2, belongs to the respiratory chain enzyme system (energy release device); ATPase Complex can be divided into three parts, and the respiratory chain can be divided into four complexes.
ATPase complex: F1-Fo coupling factor. It is a complex composed of multiple peptides. It can be divided into three parts in structure, namely the head sector, the stalk sector and the membrane sector. During the ATP synthesis process, F1 (head), OSCP (handle) and F0 factors (membrane) work together.
This part is a basic particle that protrudes from the inner membrane and is easily separated. It is called FI coupling factor, or F1 factor for short. F1 factor is an ATPase that depends on Ca2+ and Mg2+ and is composed of 10 polypeptide chains. These peptide chains are divided into five types, namely a, β, x_δ and e, with molecular weights of 56000, 52000, 32000, 21000 and 11500. Each has two molecules participating in the composition of the Fr factor. According to the enzyme activity analysis, these five polypeptides have a certain order. If these polypeptide subunits are separated from each other, they will have no enzymatic activity. However, when the a and β subunits are combined, they will exhibit ATPase activity. F1 factor binds to the handle through the delta subunit. When F1 factor binds to the handle and membrane, the ATPase becomes active and can be inhibited by oligomycin. In addition to 10 polypeptide subunits, the head also has a small heat-stable protein molecule bound to the F1 factor. This protein is called Fr inhibitory protein with a molecular weight of 10.000. Its function is to inhibit ATPase from hydrolyzing ATP, but does not inhibit ATPase from catalyzing oxidative phosphorylation.
The stalk sector is made up of a protein and is relatively easy to separate. This protein can make Fi factor sensitive to oligomycin, so that the activity of F1 in catalyzing ATP synthesis can be inhibited by oligomycin. Oligomycin cannot directly act on the FI factor, but first acts on the membrane protein component that binds to the handle, and then transmits the inhibitory effect through the handle. Therefore, the protein that constitutes the handle is called oligomycin-sensitive transfer protein, referred to as OSCP protein. Once the Fr factor is separated from the OSCP protein, it is no longer inhibited by oligomycin.
The component that makes up the membrane is called Fo factor and contains at least four types of polypeptide chains. These polypeptide chains are hydrophobic and embedded in membrane lipids. Among them, the most clearly studied is a small lipoprotein (Proteolipid), which is a non-polar substance and one of the very few proteins that can be dissolved in a variety of organic solvents. Inhibitors of ATP synthesis, such as oligomycin and N•N bicyclohexyldiimide (DCCD), can only inhibit if they bind to this lipoprotein. Therefore, lipoprotein is the oligomycin-sensitive component of Fo factor. The function of the Fo factor is related to the transfer of protons. When the F factor and OSCP are removed, the membrane becomes highly permeable to protons and protons leak into the matrix. After F and OSCP are combined on the membrane, they can block the leakage of protons. Therefore, the Fo factor may contain a channel for protons to penetrate the membrane and reach the F1 factor. Another function of Fo factor is related to the binding of Fr factor to the intima.
The electron transport chain (ETC) is an electron transport system composed of a series of electron carriers in order of gradually increasing electron affinity. All components are embedded in the inner mitochondrial membrane and are segmented into separate complexes in sequence. The physical arrangement of each carrier component within the complex also conforms to the direction of electron flow. The electron transport chain in mitochondria releases energy along with the oxidation of nutrients, also known as the respiratory chain.
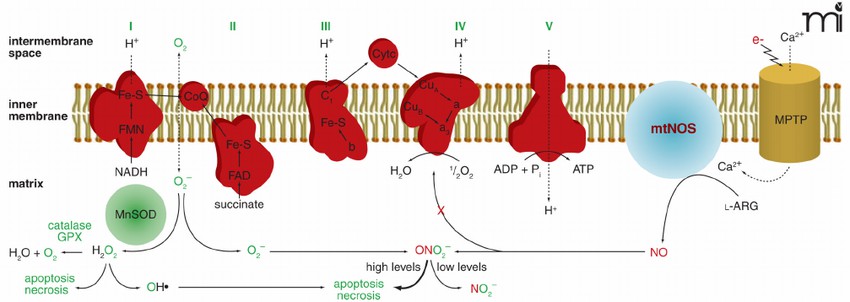 Figure 1. Schematic depicting the inner mitochondrial membrane and the five subunits of the mitochondrial electron transport chain. (Zabbarova I, et al.; 2008)
Figure 1. Schematic depicting the inner mitochondrial membrane and the five subunits of the mitochondrial electron transport chain. (Zabbarova I, et al.; 2008)
The main components of the electron transport chain in mitochondria include: 1, flavoprotein; 2, iron-sulfur protein; 3, cytochrome; 4, ubiquinone and copper atoms. They are all hydrophobic molecules with redox effects. Except for ubiquinone and copper Apart from atoms, other components are proteins, which transfer electrons through the reversible redox of their prosthetic groups. They form four complexes on the membrane surface, called complex I (NADH dehydrogenase complex), complex II (amber Acid dehydrogenase complex), complex III (cytochrome reductase complex), complex IV (cytochrome oxidase complex). NADH passes through complex I, coenzyme Q, complex III, cytochrome C, and complex IV in sequence, and finally transfers electrons to oxygen and discharges protons into the mitochondrial membrane space, and finally generates 2.5 ATP through mitochondrial ATP synthase. FADH2 is complexed Body II, coenzyme Q, complex III, cytochrome C, and complex IV finally transfer electrons to oxygen and expel protons to the mitochondrial membrane space to finally generate 1.5 ATP through mitochondrial ATP synthase. Since the amount of ATP generated by the former is greater than The latter, so the former is called the primary electron transport chain and the latter is called the secondary electron transport chain.
The main function of mitochondria is to perform oxidative phosphorylation, and under normal conditions, oxidative phosphorylation is closely coupled with electron transport. In this way, the above-mentioned components of the electron transport chain form a complete electron transport chain, which is orderly and gradually transfer electrons from reduced coenzyme to molecular oxygen. During this process, the total free energy difference between the reduced coenzyme and oxygen is divided into individual electron transfer steps, several of which are coupled to proton pumping, thus establishing and maintaining the proton migration force responsible for ATP formation.
Reference
| Target | Cat. No. | Product Name | Expression System | Tag/Conjugate | Application | |
| Mitochondrion | DAG-T1243 | Mitochondrial Antigen | Inquiry |
| Target | Cat. No. | Product Name | Size | Species Reactivity | Application | Detection Sample | |
| M2 type mitochondria | DEIA1224 | Human M2 type mitochondria Antibody ELISA Kit | 96T | Human | Quantitative | culture supernatants, serum, plasma, tissues | Inquiry |
| Mitochondria | DEIA1904 | Mitochondria ELISA Kit | 96T | Human | Qualitative | serum | Inquiry |
| ROS1 | DEIA-LL203 | Human ROS1 ELISA Kit | 96T | Human | Quantitative | Serum, plasma, tissue homogenates and other biological fluids | Inquiry |
| AMA-M2 | DEIA1826 | AMA-M2 Antibody ELISA Kit | 96T | Human | Quantitative | Serum, plasma | Inquiry |
| DEIA3129 | AMA-M2 Ab ELISA Kit | 96T | Human | Quantitative | Serum, plasma | Inquiry |