Enterovirus 71 (EV71) is a single-stranded, positive-sense RNA virus, belonging to family Picornaviridae, genus Enterovirus, species A. EV71 infection usually causes hand-foot-mouth disease (HFMD) in children under 5 years old. HFMD is usually a mild disease in children, but in some cases HFMD may be a severe neurogenic disease, complicated with aseptic meningitis, encephalitis, acute flaccid paralysis and fatal neurogenic pulmonary edema. Public health interventions to control outbreaks involve social distancing measures, but their eff ectiveness has not been fully assessed. Vaccines being developed include inactivated whole-virus, live attenuated, subviral particle, and DNA vaccines [1].
Enteroviruses are non-enveloped, spherical viruses with a diameter ranging from 28 to 30 nm, and single positive-stranded RNA. The genome of these enteroviruses is approximately 7.5-8.0 kb in length and contains one open reading frame (ORF), flanked by a highly structured 5'-untranslated region (5' UTR) and a 3' UTR with a poly(A) tail [2, 3]. The 5' UTR is composed of an RNA cloverleaf structure followed by an internal ribosomal entry site (IRES). Following entry of the virus particle into host cells and release of the viral genome from an endosome into the cytoplasm, viral RNA can be translated in an IRES-dependent manner. During translation and genome replication, the viruses require not only IRES-specific trans-acting factors (ITAFs) but also several host factors including T-cell-restricted intracellular antigen 1 (TIA-1) and TIA-1 related protein (TIAR) for effective viral replication. The interaction of TIA-1 and TIAR with the 5′ untranslated region of the viral genome can positively enhance viral replication, although ITAFs usually regulate viral growth at the translational step. The genome is initially translated into a single large polyprotein of approximately 2,200 amino acid residues. This polyprotein is proteolyzed into P1, P2, and P3 precursor proteins, and is further cleaved co- and post-translationally by viral 2A, 3C, and 3CD proteases. The P1 precursor protein is cleaved into the capsid proteins VP3, VP1, and VP0. VP0 is then further divided into VP4 and VP2. The P2 precursor protein is processed to form the viral protease 2A and the 2BC polyprotein, and the 2BC polyprotein is further cleaved into two non-structural proteins, 2B and 2C. The P3 precursor protein is initially proteolyzed into 3AB and 3CD, and then further proteolyzed to form proteins 3A, 3B, 3C, and 3D [4].
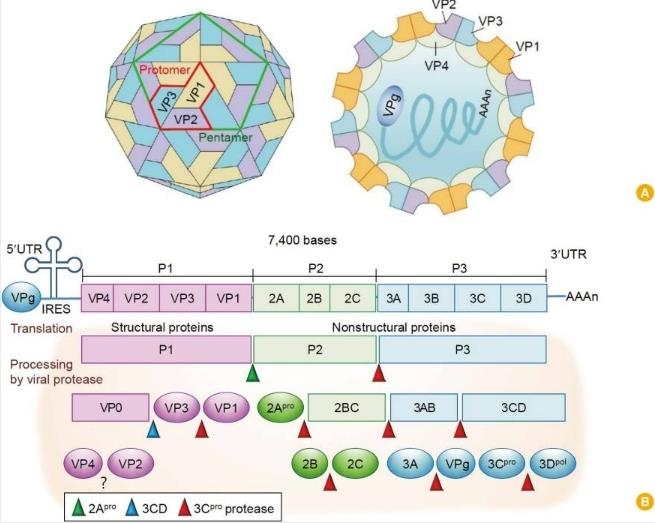 Figure 1. Structure of the enterovirus 71 (EV71) virion.
Figure 1. Structure of the enterovirus 71 (EV71) virion.
Several viral receptors that are responsible for entry of EV71 into host cells have been characterized. These receptors include human scavenger receptor B2 (hSCARB2), human P-selectin glycoprotein ligand 1 (PSGL-1), dendritic cell specific intercellular adhesion molecule-3 grabbing nonintegrin (DC-SIGN), annexin A2 (Anx2), heparan sulfate (HS), and sialylated glycan [5].
After an enterovirus binds with a specific receptor on the cell surface, a series of structural changes occur in the virus capsid (yet to be defined in EV71) and pores are formed in the cell membrane through which the virion RNA is released into the host cell cytoplasm. Being positive-sensed, the parent virus RNA acts directly as a messenger RNA and is translated into a large polypeptide that is promptly cleaved by the viral proteases into 11 mature structural and non-structural proteins. The replication of the virus genome by the error-prone RNA-dependent RNA polymerase 3Dpol takes place in a vesicle membrane structure (viral replication complex). The polymerase is estimated to misincorporate one or two bases in every genome copying event, which explains why the virus mutates and evolves rapidly. Within the VP1 gene 4.2-4.6×10-3 nucleotide substitutions occur per site per year, which is similar to the number in poliovirus and greater than that of influenza viruses [6].
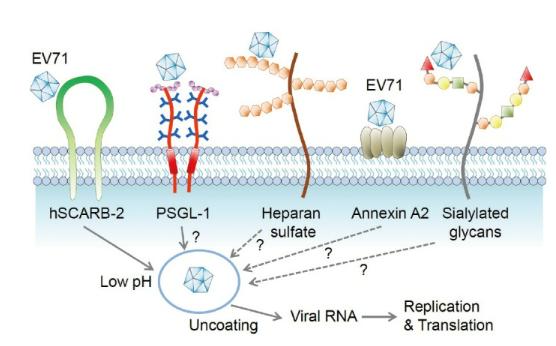 Figure 2. Viral receptors for enterovirus 71 (EV71) on host cells.
Figure 2. Viral receptors for enterovirus 71 (EV71) on host cells.
There have been several types of EV71 vaccine candidates, including attenuated strains, inactivated whole-virus, virus like particles (VLP), recombinant proteins, recombinant vectors, and peptide vaccines.
Three vaccines against EV71 have been licensed in China, all using C4 genogroup strains. The efficacy of these three vaccines after two doses immunization ranges from 90.0% to 97.4% after one year of surveillance to 95.1% after two-year follow-up. Candidate vaccines containing B4 and B5 genogroups are in development elsewhere but have not yet reached the licensing stage. Other vaccines such as recombinant and other forms of subunit vaccines, vectored vaccines, virus-like particle vaccines are in the early stage of development. In addition, the development of vaccines against Coxsackie A16, A6 and A10 is being considered with a view to developing combination/multivalent vaccines.
Immunization with recombinant VP1 protein of EV71 expressed in Escherichia coli, yeast, or the baculovirus system can induce high levels of EV71 VP1-specific IgG antibody, and confer protection against EV71 infection. Compared with the VP1 sequences of various sub-genotypes of EV71, the amino acid residues of epitope SP70 are highly conserved. Immunization with SP70 synthetic peptide, which contains a neutralizing linear epitope from the EV71 VP1 capsid protein, could elicit a neutralizing antibody titer comparable to that obtained with a whole virion-immune serum [7].
VLPs for EV71, which resemble the natural virus capsid structure, have been produced and purified as potential vaccines. Immunization with EV71 VLP was highly immunogenic and induced protective efficacy against lethal challenge in newborn mice. Based on this EV71 VLP technology, chimeric VLPs including combined SP70 epitopes of EV71 and CVA16 structural proteins or fusions of hepatitis B core antigen with SP70 epitopes of EV71 could elicit protective neutralizing antibodies in mice [8, 9].
DNA vaccines have also been tried for EV71. Immunization with DNA constructs containing the VP1 gene of EV71 could elicit the production of VP1-specific IgG and neutralizing antibodies against EV71 but showed low levels of antigenicity. Maternal immunization with an attenuated Salmonella enterica serovar Typhimurium expressing the EV71 VP1 gene conferred protection against lethal EV71 infection in the offspring. Recombinant adenovirus with the EV71 P1 and 3CD genes can enhance neutralizing antibody and protective cellular immune responses to prevent EV71 infection. Oral immunization using Tg tomato fruit expressing the EV71 VP1 protein can elicit both humoral and cellular immunity, including mucosal VP1-specific IgA antibody [10, 11].
Immunization of cynomolgus monkeys with an attenuated EV71 genotype A (BrCr) could produce high neutralization activity with cross-reactivity for other genotypes and confer protection against lethal challenge by virulent EV71 genotype A. However, this strategy needs to overcome some safety issues, since the attenuated strain itself caused mild neurological symptoms and was still neurotropic when inoculated via the intravenous route. A high-fidelity variant of EV71 with the two amino acid modifications, L123F and G64R, in the viral 3D RNA polymerase exhibited an attenuated phenotype and showed potential as a live attenuated EV71 vaccine [12, 13].
Among the various vaccine candidates, inactivated whole virus vaccines are the preparation of choice capable of fulfilling the demand for effective control. Development of inactivated whole-virus EV71 vaccines has progressed rapidly, inspired by previous developments in inactivated vaccines. Immunization with formalin inactivated EV71 strain can elicit high levels of virus-specific antibody including cross-neutralizing activity and protects the immunized host against lethal challenge with virulent EV71 in the murine model. Based on successful pre-clinical work, phase I or phase II clinical trials of candidate inactivated EV71 vaccines were conducted. Vaccination induced significantly greater neutralizing antibody and specific T-cell responses in vaccinees without a marked inflammatory response. Other inactivated EV71 vaccine candidates can elicit cross-neutralizing antibody responses against EV71 sub-genotypes B1, B4, B5, and C4A [14, 15].
References
| Cat. No. | Product Name | Species Reactivity | Sample | |
| DEIA081 | EV71 IgM ELISA Kit | Human | serum, plasma | Inquiry |
| DEIA435 | EV71 IgM DOT-ELISA Kit | Human | serum, plasma | Inquiry |
| DEIA3406 | EV71 VP1 ELISA Kit | Human | serum, plasma | Inquiry |
| DEIA-XY120 | Enterovirus IgA ELISA Kit | Human | serum, EDTA plasma, heparin plasma, citrate plasma | Inquiry |
| DEIA-XY121 | Enterovirus IgG ELISA Kit | Human | serum, EDTA plasma, heparin plasma, citrate plasma | Inquiry |
| DEIA-XY122 | Enterovirus IgM ELISA Kit | Human | serum, EDTA plasma, heparin plasma, citrate plasma | Inquiry |
