Diphtheria is caused by bacterial exotoxins secreted by toxigenic bacteria of the Corynebacterium diphtheriae or the rare Corynebacterium ulcerans, and can lead to respiratory diphtheria and cutaneous diphtheria, with respiratory diphtheria having a higher risk of death. Infection usually develops from prodromal symptoms to membranous inflammation of the pharynx, tonsil, or throat. Diphtheria toxin can spread throughout the body, resulting in irreversible inhibition of protein synthesis in the respiratory system, kidney, heart and nervous system, and epithelial cell dysfunction.
Diphtheria is common in children under the age of 12, and newborns have a higher risk of infection due to lack of immunity. The mortality rate for untreated patients is between 5% and 20%.
Diphtheria toxin (DT) is known to be the main virulence factor of Corynebacterium diphtheriae. It is an exotoxin that can be secreted through the plasma membrane without causing cell lysis. The mature exotoxin is a 58kD peptide consisting of 535 amino acid residues. Under the action of mild trypsin in vitro, DT is reduced to two fragments, A and B, which maintain covalent connection through interchain disulfide bonds. Fragment A has enzyme activity, while fragment B binds the protein to the cell surface receptor and promotes the translocation of fragment A to the cytoplasm. DT consists of three domains: catalytic domain (C) corresponding fragment A (21kD), transmembrane domain (T) (20kD) and receptor binding domain (R) (17kD) to form fragment B (37kD).
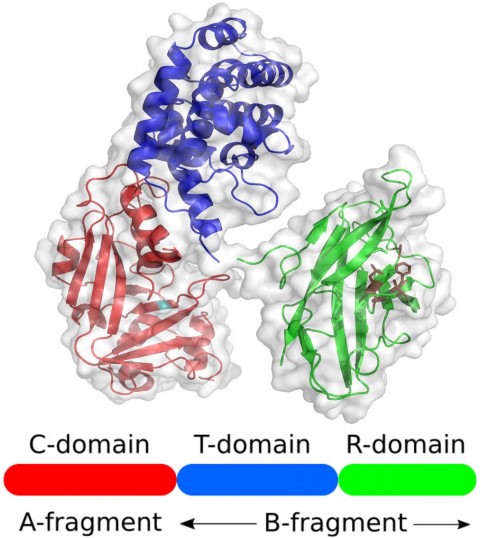 Figure 1. Protein structure of diphtheria toxin
Figure 1. Protein structure of diphtheria toxin
(Source: Wenzel EV, et al. 2020)
DT is endocytosed into the cell through the binding of its B subunit to heparin-binding epidermal growth factor-like growth factor (HB-EGF); the ADP-ribosylation activity of DT prevents protein synthesis in the host cell, leading to cell death.
Diphtheria toxin inhibitor (DtxR) is a divalent metal-activated transcriptional inhibitor that regulates siderophore-mediated iron uptake and heme oxygenase and DT expression. The functional activity of DtxR is controlled by iron. The binding of Fe2+ leads to the isomerization of DtxR conformation, which is beneficial to the binding of activation inhibitor to tox operator. In silico analysis, it was found that DtxR may provide molecular bonds for Fe2+-induced Fenton effect (production of reactive oxygen species harmful to bacteria) to protect bacterial DNA from oxidative damage. Gene interference analysis shows that DtxR has a great impact on the expression of many genes, even under a high iron environment, the gene inactivation of DtxR will lead to the expression of constitutive DT.
 Figure 2. Mechanism of action of diphtheria toxin
Figure 2. Mechanism of action of diphtheria toxin
(Source: Sharma NC, et al. 2019)
The use of diphtheria toxoid vaccines, especially the three-dose series containing diphtheria-tetanus-pertussis (DTP), has resulted in a significant reduction in diphtheria cases. It can effectively avoid the occurrence of severe illness or even death, but has little preventive effect on mild infection and asymptomatic infection. Past studies based on our understanding of the dynamics of the diphtheria epidemic has made inappropriate assumptions, including that vaccination gains colonization immunity, that immunity is lifelong and does not wane, and that only symptomatic people are contagious, which has led to flaws in the measures we have taken to control the disease.
Diphtheria toxoid vaccines prevent diseases by stimulating the production of antitoxin antibodies, protecting 90% of clinical diphtheria and 50% of complications of diseases such as myocarditis, neurological impairment, and death. Diphtheria toxoid vaccine can reduce the risk of severe illness and death, full vaccination can effectively prevent 81% of severe cases and 93% of deaths, and partial vaccination can effectively prevent 47% of severe cases and 68% of deaths.
Table 1. Relative risk of mortality among diphtheria cases, by age, vaccination status, and antitoxin receipt
| Characteristic | Relative Risk of Mortality (95% Crl) |
| Age, y | |
| 0-4 | 1.5 (1.4-1.6) |
| 5-19 | 0.80 (.76-.89) |
| ≥20 | Reference |
| Vaccination | |
| None | Reference |
| Partial (1-2 doses) | 0.32 (.23-.44) |
| Full (≥3 doses) | 0.07 (.04-.10) |
| Antitoxin | |
| No | Reference |
| Yes | 0.24 (.22-.28) |
(Source: Truelove SA, et al. 2020)
The main vaccination series of DTP include doses for 6, 10 and 14 weeks, and enhanced doses for 12-23 months, 4-7 years, 9-15 years, and adults every 10 years. A meta-analysis of diphtheria cases and vaccine efficacy showed that the effective rate of complete vaccination (≥ 3 doses) was 87% and that of incomplete vaccination was 71% (1-2 doses).
DTP has good safety and low immunogenicity. Clinical trials found that adults treated with DTP had no increased risk of adverse events compared with adults who received DTP 10 years after the first dose of DTP. Few adverse reactions were reported in pregnant women who received multiple doses of DTP during one pregnancy and those who received additional doses of vaccine. The most common adverse reactions were pain at the injection site, fatigue, and headache.
The immunity brought by the vaccine gradually weakens over time. In the population with low enhanced vaccination rate, the effective rate of full vaccination against symptomatic diseases is 96% in children aged 0-4 years old, 92% in children aged 5-19 years old, and 63% in children aged ≥ 20 years old. Serological studies have also shown that immunity is weakening, and the proportion of individuals with fully protective antibody levels (≥ 0.1 IU/mL) has decreased by 0.6% a year since vaccination, consistent with the decline in vaccine effectiveness.
Diphtheria traditionally combines with tetanus and pertussis antigens to produce diphtheria-tetanus-pertussis vaccine, but now it often binds to other antigens, such as Haemophilus influenzae B (Hib), hepatitis B and polio inactivated vaccines.
The epidemiology of C. diphtheriae has changed. In some countries with high coverage of anti-diphtheria vaccination, there are more and more cases of infection caused by non-toxic diphtheria strains, a large proportion of which are invasive infections, including bacteremia, septicemia, and endocarditis. Therefore, a new vaccine is needed, which can effectively prevent not only diphtheria but also infection caused by non-virulent strains.
There are still some research difficulties in improving the efficacy of the vaccine, including the lack of international standards for diphtheria toxoid used in the flocculation test (a test that binds antigens and antibodies to measure anti-diphtheria-tetanus antibody levels), and the concentration of heavy metals in the blood may lead to a decrease in vaccine-induced antibody levels. Some studies have found that diphtheria vaccination combined with vitamin A injection will produce a strong antibody response. In addition, fully vaccinated people are still likely to be infected with diphtheria and develop typical symptoms, possibly as a result of infection with non-toxigenic Corynebacterium that is not protected by the vaccine.
We should continuously monitor changes in pathogens, improve existing vaccines or develop new vaccines that can induce effective mucosal defence in order to provide early and lasting protection for infants, adults and the elderly in response to changing epidemic situations.
References
| Target | Cat. No. | Product Name | Size | Species Reactivity | Application | Detection Sample | |
| Diphtheriae | DEIA05524 | Corynebacterium diphtheriae IgG - ELISA Kit | 96T | Human | Quantitative | Serum, citrate plasma | Inquiry |
| Target | Cat. No. | Product Name | Expression System | Tag/Conjugate | Application | |
| C. diphtheriae | DAGC697 | Native C. diphtheriae Antigen | N/A | Unconjugated | Immunogen, WB, ELISA | Inquiry |
| Target | Cat. No. | Product Name | Host | Isotype | Application | |
| C. diphtheriae | DPAB-CS24018G | Human Anti-C. diphtheriae IgG Control Serum | Human | IgG | ELISA | Inquiry |
| Target | Cat. No. | Product Name | Size | Species Reactivity | Application | Detection Sample | |
| Diphtheria toxin | DEIA1776 | Human Anti-Diphtheria Toxoid IgG ELISA Kit | 96T | Human | Quantitative | Serum or plasma | Inquiry |
| Target | Cat. No. | Product Name | Expression System | Tag/Conjugate | Application | |
| C. diphtheriae toxin | DAGH042 | C. diphtheriae toxin | N/A | Unconjugated | N/A | Inquiry |
| DAG4060 | Recombinant C. diphtheriae Toxin (mutation CRM197) | P. fluorescens | Unconjugated | N/A | Inquiry |
| Target | Cat. No. | Product Name | Size | Species Reactivity | Application | Detection Sample | |
| Diphtheria Toxoid | DEIASL262 | Mouse Anti-Diptheria Toxoid IgG ELISA Kit | 96T | Quantitative | Serum | Inquiry | |
| DEIASL270 | Rabbit Anti-Diptheria Toxoid IgG ELISA Kit | 96T | Quantitative | Serum | Inquiry | ||
| DEIA328 | Diphtheria Toxoid IgG ELISA Kit | 96T | Human | Quantitative | Serum or plasma | Inquiry | |
| DEIAJX005 | Human Anti-Diphtheria Toxin/Toxoid (DTX) IgA ELISA kit | 96T | Quantitative | Serum and other biological fluids | Inquiry | ||
| DEIAJX002 | VacciGel Diphtheria Toxoid ELISA Kit | 96T | Quantitative | Vaccine formulated in Alum | Inquiry | ||
| DEIAJX003 | Monkey Anti-Diphtheria Toxin/Toxoid (CRM197) IgG ELISA kit | 96T | Quantitative | Serum or plasma samples | Inquiry |
| Target | Cat. No. | Product Name | Expression System | Tag/Conjugate | Application | |
| C. diphtheriae Diphtheria Toxoid | DAG2689 | C. diphtheriae Diphtheria Toxoid | N/A | Unconjugated | ELISA | Inquiry |