Nipah virus (NiV) in the family Paramyxoviridae is capable of causing zoonotic diseases, with severe respiratory and neurological manifestations in humans. More specifically, NiV belongs to the genus Henipavirus. Different strains of the virus cause different morbidity and mortality rates, with up to 70% of patients dying, which has led to the classification of NiV as a 4 groups of risk pathogens that can only be studied in a Biosafety Level 4 (BSL-4) facility.
NiV is a negative single-stranded RNA virus with full-length 18kb, in which the P gene encodes non-structural proteins C, V, and W, which plays a key role in the pathogenesis and anti-innate immune response. NiV contains proteins similar to other paramyxoviruses, including nucleocapsid protein (N), phosphoprotein (P), matrix protein (M), glycoprotein F (F), glycoprotein G (G), and ribonucleic acid polymerase. Glycoproteins G (G) and F (F) are essential for virus-host cell binding, fusion, and germination. Unlike other paramyxoviruses, NiV contains reticular cytoplasmic inclusions and has no hemagglutinin and neuraminidase characteristics.
The replication and transcription of NiV requires that the antisense RNA binds closely to the N protein and RNA polymerase complex. In the process of transcription, the RNA polymerase complex first replicates viral RNA, and then produces capped RNA, short uncapped RNA, and polyadenylate mRNA that encode proteins.
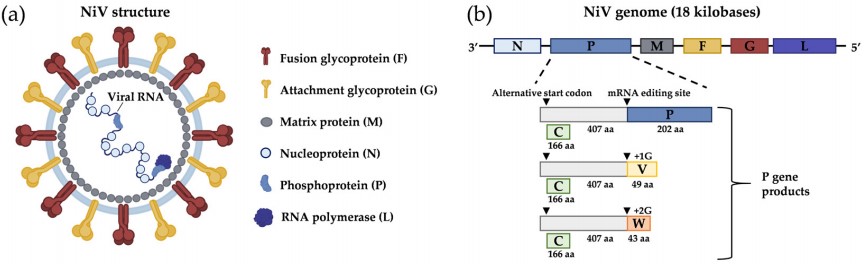 Figure 1. Nipah virus structure and genome organization
Figure 1. Nipah virus structure and genome organization
(Source: Garbuglia AR, et al. 2023)
NiV's host in nature is Pteropus fruit, and infected bats are able to transmit NiV viruses through saliva, urine, sperm, and feces. Because of their adaptively enhanced immune tolerance, bats are reservoirs of asymptomatically carried viruses. Serological studies have found that Pteropus fruit from different countries have reactivity and antibodies that neutralize NiV, which can cause fatal infections in humans.
The virus was first identified in Malaysia, where cases of fever, headache, and loss of consciousness were reported in September 1998 in what was initially thought to be an outbreak of Japanese encephalitis, but control measures did not prevent the outbreak. In March 1999, a new virus was isolated from the cerebrospinal fluid of a patient in the village of Nipah, Sungai, giving it the name Nipah virus. The outbreak resulted in 283 symptomatic infections and 109 deaths. Epidemiology of the outbreak revealed that pigs and pig feces were risk factors for transmission. Pigs became infected with NiV through the use of fruit bitten by bats, thereby transmitting the virus to other animals and humans.
The epidemiology of NiV in Bangladesh has begun to change, with the addition of transmission in the form of consumption of raw palm sap, in addition to contact with pigs, which is the most common mode of transmission in Bangladesh. Since 2001, there have been seasonal outbreaks of NiV infections in Bangladesh, timed to coincide with sap harvesting seasons (December to May). The absence of large slaughterhouses in Bangladesh makes animal-to-animal transmission unlikely, despite the presence of very high seroprevalence of pigs for NiV. In contrast, human-to-human transmission is very important in the spread of outbreaks in Bangladesh, with a large human-to-human outbreak in Faridpur in 2004.
India had two outbreaks of NiV in the border areas of Bangladesh with 66 suspected cases and 45 deaths in 2001 and 5 cases and 100% deaths in 2007. Both outbreaks were considered for reasons of proximity to the Bangladeshi outbreak area, however, in 2018 there was an outbreak of NiV in a geographically isolated area from the outbreak area. Nosocomial second infections were present in the Indian outbreak.
In 2014, NiV caused 17 infections in the Philippines and 82% of patients died. Ten of these patients had an epidemiologic history of close contact with horses or consumption of horse meat, and 10 horses died during the same period, nine of which showed neurologic symptoms. There were also healthcare workers who were infected with the virus due to human-to-human transmission. The strain circulating in the Philippines is closely related to the Malaysian strain, but no clear human-to-human transmission of the previous Malaysian strain has been found, suggesting the possibility of co-evolution of the different NiV strains, as well as the possibility of strain mutation.
Table 1. Summary of some NiV outbreak data
| Region and/or country | Month and year | No. of cases | No. of deaths | Case fatality (%) |
| Malaysia | September 1998-December 1999 | 265 | 105 | 39.6 |
| Singapore | March 1999 | 11 | 1 | 9.1 |
| Siliguri (India) | April-May 2001 | 66 | 45 | 68.2 |
| Rajbari, Faridpur (Bangladesh) | January-April 2004 | 67 | 50 | 74.6 |
| Tangail (Bangladesh) | January-March 2005 | 12 | 11 | 91.7 |
| Nadia (India) | April 2007 | 5 | 5 | 100 |
| Comilla, Nilphamari, and Rangpur (Bangladesh) | January-February 2011 | 44 | 40 | 90.9 |
| Kozhikode, Malappuram (India) | May 2018 | 18 | 17 | 94.4 |
(Source: Askari M, et al. 2023)
Human infection with NiV does not result in immediate illness, with an incubation period of approximately 4 to 21 days, after which prodromal signs and symptoms appear, including fever, headache, and myalgia. A small proportion of infected individuals have no obvious symptoms. NiV causes acute encephalitis and respiratory illness, with encephalitis manifesting within a week of onset, often with altered mental status, hypotonia, segmental myoclonus, and gaze paralysis. The disease is rapidly progressive and has a high mortality rate. Residual neurological deficits such as fatigue, focal neurological deficits, and depression occur in 20% of survivors.
The clinical features of infection vary between countries. In India and Bangladesh, 70% of patients present with respiratory illness (cough, dyspnea, atypical pneumonia), while in Malaysia neurological symptoms are more common. In addition, the health status of the infected person influences the clinical prognosis. Risk factors such as advanced age, comorbidities, thrombocytopenia, and elevated aminotransferases on admission and seizures increase the poor prognosis of patients.
Early diagnosis of suspected cases can quickly limit the spread of the epidemic and provide patients with appropriate medical treatment. NiV detection methods can be categorized into direct and indirect detection. Direct detection refers to the NiV itself, such as nucleic acid amplification and gene sequencing for direct detection of the NiV genome, and immunohistochemistry or immunofluorescence for detection of NiV surface proteins. Indirect tests assess NiV infection by detecting anti-NiV antibodies, such as enzyme-linked immunosorbent assay (ELISA). Currently, real-time quantitative PCR is the best choice for detecting NiV, as it has high sensitivity and specificity and can provide timely results in a relatively short time. On the other hand, the method applies to a wide range of sample types, including nasal swabs, cerebrospinal fluid, urine, and blood. The Centers for Disease Control and Prevention has developed routine PCR assays targeting the nuclear envelope protein gene. A TaqMan probe-based assay developed in 2004 detects the N gene with a very high sensitivity of 1 pfu, is specific for NiV RNA, and can be used for outbreak diagnosis. A SYBR Green-based assay for different regions of the N gene has also been developed, which has a lower sensitivity (100 pfu) but can also detect HeV.
Table 2. Molecular diagnostic assay of Nipah virus
| Test type | Virus detected | Target gene | Sensitivity | Specificity |
| Real-time RT-PCR with fluorescent reporter dye detected at each PCR cycle | NiV | N gene of NiV M strain | Whole blood: 103 copies/mL | No cross-reactivity with measles virus was declared |
| SYBR Green RT-PCR | NiV | N gene | 20 copies/reaction | Whole blood: 100% for Nipah virus |
| Transcription-loop-mediated isothermal amplification (RT-LAMP) | NiV | N gene of NiV | 107 copies/reaction | No cross reactivity with HeV, Newcastle disease virus, Japanese encephalitis virus, and Influenza A virus |
| NGS | 35 epizootic and zoonotic viruses | Full genome | 21 genomes/reaction | ND |
| Real-time TaqMan RT-PCR with fluorescent reporter dye detected at each PCR cycle | Several pathogens | N gene | Whole blood: 54 copies/well | 100%; no cross-reactivity with other Viral Haemorrhagic Fever |
(Source: Garbuglia AR, et al. 2023)
Serologic testing is indicated for epidemiologic investigations and outbreak surveillance and may help detect NiV. IgM and IgG antibodies typically appear during the first week of a patient's illness in the serum and cerebrospinal fluid (CSF). They can persist in the patient for long periods, with 50% of patients being 100% IgM positive three days after symptom onset and 31% being 100% IgG positive on day 17. Ongoing follow-up of NiV-positive patients has shown that IgM is present in symptomatic patients as late as day 27, while IgG persists for more than a year. In a small study conducted in India, contact cases of NiV-positive patients were tested periodically over several months for IgM and IgG antibodies. Antibodies were detected in three symptomatic and two asymptomatic contact cases. In symptomatic patients, IgM was detectable from days 5 through 27 after the onset of symptoms, while IgG persisted for more than a year. Similar IgM and IgG immune responses to NiV infection were observed with and without clinical symptoms.
References
| Target | Cat. No. | Product Name | Host | Isotype | Application | |
| HeV and NiV Glycoprotein | CABT-NS1156 | Human Anti-HeV and NiV Glycoprotein Monoclonal Antibody, clone 48D3 | Human | IgG | IP, WB | Inquiry |
| NiV F F1 | CABT-NS1194 | Mouse Anti-NiV F F1 Monoclonal Antibody, clone 5G7 | Mouse | IgG2b | IP, WB | Inquiry |
| NiV F F2 | CABT-NS1195 | Mouse Anti-NiV F F2 Monoclonal Antibody, clone 6D3 | Mouse | IgG1 | IP, WB | Inquiry |
| NiV F Site I | CABT-NS1197 | Mouse Anti-NiV F Site I Monoclonal Antibody, clone 16C2 | Mouse | IgG1 | IP | Inquiry |
| NiV F Site II | CABT-NS1192 | Mouse Anti-NiV F Site II Monoclonal Antibody, clone 4C9 | Mouse | IgG1 | IP | Inquiry |
| NIV G Protein | DMAB-A001 | Mouse Anti-Nipah Virus G Protein monoclonal antibody, clone D12085N | Mouse | IgG1 | ELISA | Inquiry |
| NIV gF | CABT-L1230M | Mouse Anti-Nipah Virus Glycoprotein F monoclonal antibody, clone DH22 | Mouse | IgG | WB, ELISA | Inquiry |
| Target | Cat. No. | Product Name | Expression System | Tag/Conjugate | Application | |
| Nipah Virus | DAG-WT1059 | Recombinant Nipah Virus Glycoprotein G | Insect cells | Unconjugated | Immunoassays | Inquiry |
| Nipah Virus | DAG-WT633 | Recombinant Nipah Virus F Protein [His] | HEK293 | His | ELISA | Inquiry |
| Nipah Virus | DAG-WT634 | Recombinant Nipah Virus G Protein [His] | HEK293 | His | ELISA | Inquiry |
| Nipah Virus | DAG-WT272 | Recombinant Nipah Virus F Protein [Fc] | HEK 293 | Fc | SDS-PAGE | Inquiry |
| Nipah Virus | DAG-WT273 | Recombinant Nipah Virus Nucleoprotein [His] | HEK 293 | His | ELISA | Inquiry |
| Nipah Virus | DAG-WT381 | Recombinant Nipah Virus (NiV) VLP | HEK293 | Unconjugated | Immunoassays | Inquiry |