 Fig. 1 Structure of Zika virus
Fig. 1 Structure of Zika virus
(Sirohi D, Kuhn RJ. J Infect Dis. 2017)
Zika virus is a mosquitoborne flavivirus that, upon infection, results in teratogenic effects and neurological disorders. The emergence of Zika virus as a major public health threat has focused research on understanding virus biology and developing a suite of strategies for disease intervention. Neither an effective treatment nor a vaccine is available for Zika virus; therefore, the public health response primarily focuses on preventing infection. Different platforms have been evaluated for the design of vaccines, including DNA, mRNA, peptide, protein, viral vectors, virus-like particles (VLPSs), inactivated-virus, and live-attenuated virus vaccines and most of these vaccines have entered clinical trials.
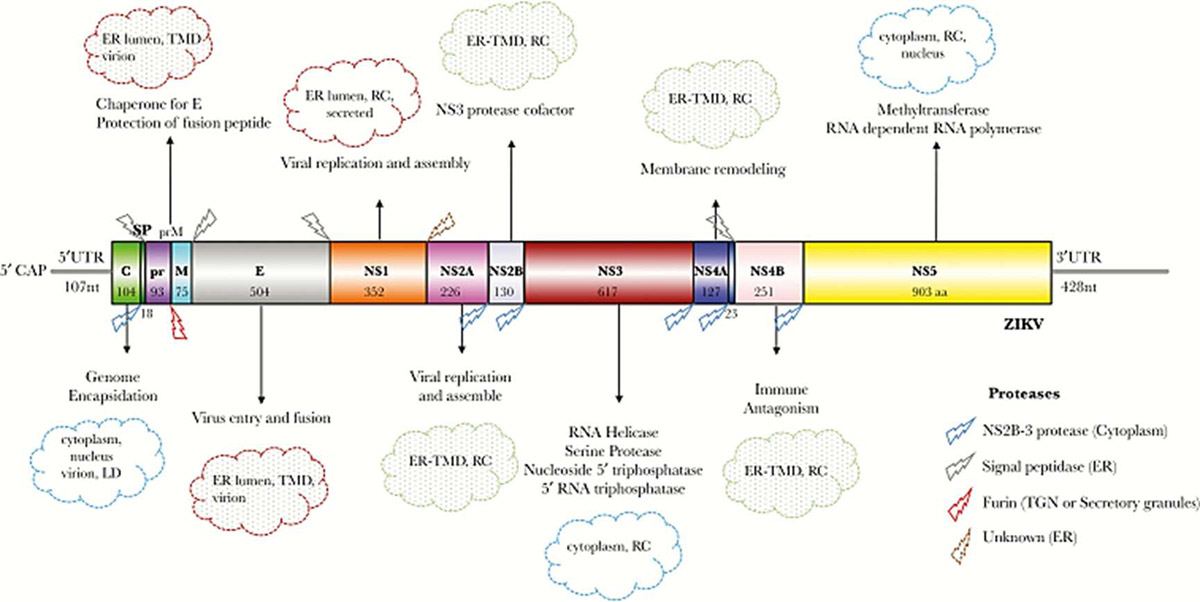 Fig. 2 The genome of Zika virus
Fig. 2 The genome of Zika virus
(Sirohi D, Kuhn RJ. J Infect Dis. 2017)
Zika virus belongs to the Flaviviridae family. It is a spherical, enveloped virus with icosahedral symmetry. The diameter of the mature virus is approximately 50 nm, and the diameter of the immature virus is closer to 60 nm. Zika virus is a positive sense, single-strand ribonucleic acid (RNA) virus with a genome size of approximately 10.8 kilobases. The RNA is translated into a single polyprotein (3423 amino acids in length) encoding 3 structural proteins—capsid (C); membrane (M), which is generated from its precursor premembrane (prM); and envelope (E)—as well as 7 nonstructural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5).
The structural proteins are essential for genomic replication and host immunity. Glycoprotein E is a major target for antibodies as it is responsible for facilitating viral entry into the host. Zika virus is capable of mediating the antibody response by the changing amino acids that surround the Asn154 glycosylation site in the virus E protein. The function of PrM proteins affects the infectivity and pathogenicity of viruses. The capsid protein has been shown to play an important role in viral assembly. NS proteins play a major role in evading host immune responses specifically by interfering with the interferon (IFN) I response.
The first report of the structure of a broadly neutralizing murine antibody (2A10G6) Fab bound to the ZIKV E protein using x-ray crystallography revealed binding to the flavivirus highly conserved fusion loop. It is expected that binding to the fusion loop will prevent the E protein from engaging with the membrane as required for fusion. Another structural study shows that the human monoclonal (C10) which was originally isolated from a DENV-infected patient binds to all 180 E proteins. The Fab binds at both ends of the E protein dimer. This antibody neutralizes virus infectivity by cross-linking the E proteins and preventing the required prefusion conformational changes. ZIKV-117, A ZIKV-specific monoclonal antibody isolated from an infected patient was shown to be a highly potent neutralizing antibody with no cross-reactivity against other flaviviruses. This antibody cross-links monomers within the dimer as well as between neighboring dimers that locks E proteins together, preventing the reorganization required for membrane fusion and virus infection.
Currently, multiple vaccine platforms are being incorporated to create new vaccines, such as DNA, mRNA, peptide, protein, viral vectors, virus-like particles (VLPs), inactivated-virus, and live-attenuated virus.
Whole inactivated virus vaccines have been effectively used for other flaviviruses including YFV, JEV, and TBEV. For ZIKV, currently, 5 of 7 inactivated virus vaccines have completed phase 1 clinical trials. Purified formalin-inactivated ZIKV vaccine (ZPIV; derived from Puerto Rico strain PRVABC59) was shown to protect against ZIKV infection in mice and non-human primates. In clinical phase I trials, ZPIV caused only mild to moderate adverse events and has been demonstrated to be well-tolerated and immunogenic. In all the vaccines, formalin was used to inactivate the virus.
Two approaches have been taken to generate live-attenuated ZIKV vaccines: (1) Engineering attenuating mutations in an authentic ZIKV isolate or (2) generating a chimeric flavivirus using DENV, JEV, or YFV to express ZIKV prM-E genes.
For (1), Shan et al. developed a vaccine using an infectious cDNA clone with 10 nucleotide deletions within the 3`untranslated region of the viral genome. This attenuated vaccine induced sterilizing protection in A129 mice (IFN-α/β receptor knockout) and NHPs. Chimeric vaccines are another version of live-attenuated vaccines. For (2), Xie et al. used a reverse genetics approach to develop two chimeric viruses by swapping the prM-E (premembrane and envelope) structural genes between DENV-2 and ZIKV, incorporating a full-length cDNA clone. These chimeric viruses also protected mice against DENV-2 and ZIKV challenge and can be developed into effective vaccines against ZIKV. In addition, the NIAID is developing a chimeric ZIKV vaccine that expresses the ZIKV prM-E genes in the context of an attenuated DENV2 backbone.
Subunit vaccines express two virion surface glycoproteins of prM and E using DNA, mRNA, or viral vectors. Alternatively, recombinant E protein or purified virus-like particles (formed by prM and E proteins) could be used directly as subunit vaccines.
Gaudinski et al. tested two DNA vaccine candidates: plasmid VRC5288 and VRC5283. VRC5283, which encodes the wild-type ZIKV sequence, showed higher immunogenicity and was more efficient thanVRC5288. Currently, VRC5283 is in Phase 2 investigation undergoing clinical trials. Yebas et al. tested a DNA subunit vaccine (GLS-5700) containing a consensus sequence of prM-E from pre-2016 human ZIKV strains, and found no serious adverse effects.
Themis Biosciences produced a recombinant measles vector that expresses the prM-E proteins of ZIKV virus and started phase 1 clinical evaluation. Preclinicial studies of a rhesus adenovirus serotype 52 vaccine that expresses the M and E proteins from ZIKV showed protection from 2 strains of ZIKV with only a single dose. Additional approaches using an MVA vector expressing NS1 (Geovax) and a VSV vector expressing prM-E (Harvard) are also in development.
Another approach to ZIKV vaccination uses an in vitro produced viruslike particle or protein subunits. Several forms of ZIKV VLPs have been reported featuring the co-expression of the prM-E, prM-E-NS1, C-prM-E, and NS2B/NS3 viral genes in human cells. The ZIKV VLPs developed by NewLink Genetics (USA) and CEVEC Pharmaceuticals (Germany) were produced by ZIKV prM-E over-expression in human CAP cells. The ZIKV VLPs developed by VBI Vaccines (USA) are retroviral GAG-derived enveloped VLPs produced by ZIKV E-NS1 expression in HEK 293 cells. Due to the E dimer has been defined as the antigenic target for neutralizing antibodies, there are also some efforts to use soluble E proteins as vaccine antigens. Peptide vaccines, designed to induce antibodies to specific epitopes, are also being explored in preclinical studies.
RNA vaccines have an added advantage over DNA vaccines in that they have no risk of possible integration into the human genome. Pardi et al. developed a ZIKV vaccine in which the prM-E proteins were encoded by mRNA. The vaccine-elicited antibodies that protected C57BL/6 mice from the ZIKV H/PF/2013 challenge. Chahal et al. designed an RNA vaccine candidate expressing prM-E using the Asian ZIKV isolate Z1106033. Immunization through intramuscular injection elicited ZIKV E protein-specific IgG responses and protected C57BL/6 mice.
A number of challenges remain to be addressed for ZIKV vaccine development. Firstly, since ZIKV, DENV, and other flaviviruses co-circulate in many geographic regions, such cross-reactive antibody-mediated enhancement poses a major challenge for ZIKV vaccine development. Secondly, if viral antigens are responsible for the cause of GBS disease, we need to identify the antigens that elicit antibodies that attack the peripheral nerves. Once such viral antigens have been defined, the vaccine candidates should be re-engineered to exclude the cross-reactive epitopes. Thirdly, ZIKV vaccine must be safe in women of childbearing age and pregnant women to avoid complications. Fourthly, since antibodies against viral prM and E proteins cross-react among different flaviviruses, it is critical to develop a serologic assay that could differentiate among pre-existing flavivirus antibodies, ZIKV vaccine-induced antibodies, and subsequent ZIKV infection-induced antibodies. Such virus-type-specific serologic assay will greatly facilitate vaccine efficacy trials.
References
| Cat. No | Product Name | Expression System | Application | |
| DAGA-3054 | Native ZIKV antigen (Strain MR 766) | E6 cells | ELISA | Inquiry |
| DAG-WT303 | Recombinant ZIKV VLP (E, pre-M, M) | HEK293 cells | WB, ELISA | Inquiry |
| DAGA6128 | Recombinant ZIKV E protein [His] | Insect cells | LFIA, ELISA | Inquiry |
| DAGA-038 | Recombinant ZIKV NS1 Protein [His] | Insect cells | LFIA, ELISA | Inquiry |
| DAGA-310 | Recombinant ZIKV Pre-M (Brazil) [His] | E. coli | WB, ELISA | Inquiry |
| DAGA-311 | Recombinant ZIKV Capsid (Brazil) [His] | E. coli | WB, ELISA | Inquiry |
| DAGA-312 | Recombinant ZIKV Capsid (Uganda) [His] | E. coli | WB, ELISA | Inquiry |
| DAGA-313 | Recombinant ZIKV E (domain III) [His] | E. coli | WB, ELISA | Inquiry |
| Cat. No | Product Name | Species Reactivity | Sample | |
| DEIABL43 | Anti-ZIKV IgM ELISA Kit | Human | Serum, plasma | Inquiry |
| DEIAJX001 | Anti-ZIKV NS1 IgG ELSIA Kit | Human | Serum and other biological fluids | Inquiry |