
Liver cancer is the fifth most common cancer in the world and the second leading cause of cancer death. Cholangiocarcinoma is the second most common liver cancer. It is a malignant tumor of bile duct epithelium. It is clinically asymptomatic and the global incidence is increasing. Due to the lack of early markers for diagnosis, most patients with cholangiocarcinoma are identified as advanced and metastatic, and eventually die.
Existing studies have found that elevated bile acid (BA) can cause liver cell inflammation, apoptosis and liver cell necrosis, so long-term elevated BA levels in patients are considered to be risk factors for the development of liver cancer. In fact, changes in serum BA may be more useful in the diagnosis of cholangiocarcinoma. Therefore, as a key factor that mediates bile acid homeostasis, JNK has been focused on research. Previous studies have determined that liver c-Jun NH2 terminal kinase (JNK) deficiency can inhibit cholangiocarcinoma cell proliferation and oncogenic transformation in a p53/Kras-induced cholangiosarcoma model; and it is deficient in hepatocellular carcinoma with diethylnitrosamine and NEMO promotes cholangiocarcinoma in the syndrome model. Interestingly, recent studies have found that liver JNK alone is not enough to develop cholangiocarcinoma.
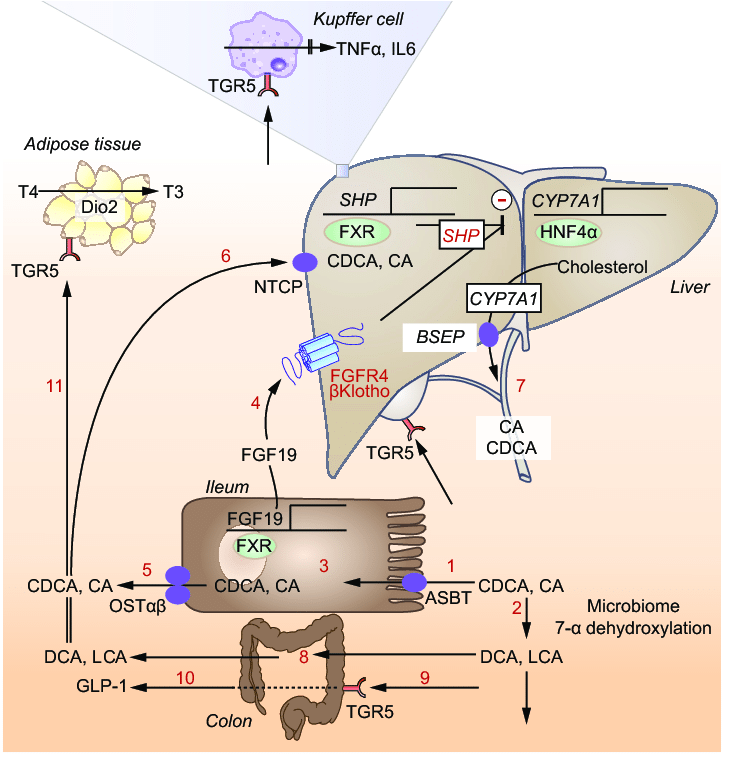
Through the search, scientists discovered an important regulator involved in lipid and BA liver metabolic homeostasis, the receptor α (PPARα) activated by peroxisome proliferators. The study found that JNK can inhibit the activation of PPARα in liver cells, thereby affecting lipid metabolism and steatosis through the liver factor FGF21 (encoded by the PPARα target gene). Further research found that JNK-mediated inhibition of PPARα leads to changes in BA homeostasis, thereby inhibiting cholangiocyte proliferation. Therefore, JNK deficiency will stimulate the proliferation of cholangiocarcinoma cells and promote the development of cholangiocarcinoma. Increased proliferation is caused by changes in BA metabolism and increased liver expression of FXR / FGF15 / FGFR4, which triggers ERK activation in cholangiocellular cells, which in turn activates downstream proliferation-related genes. The activation of FXR by BA triggers human fibroblast growth Factor 15/19 (FGF15 / FGF19) secretion. Clinical studies have found that FGF15/19 is expressed in the liver during the development of hepatocellular carcinoma and intrahepatic cholangiocarcinoma. It is worth noting that FGF15/19 improves blood glucose response and reduces hepatic steatosis, while also promoting liver cancer. Therefore, the steady state change of BA causes FGF15/FGF19 to promote the development of cholangiocarcinoma.Through the search, scientists discovered an important regulator involved in lipid and BA liver metabolic homeostasis, the receptor α (PPARα) activated by peroxisome proliferators. The study found that JNK can inhibit the activation of PPARα in liver cells, thereby affecting lipid metabolism and steatosis through the liver factor FGF21 (encoded by the PPARα target gene). Further research found that JNK-mediated inhibition of PPARα leads to changes in BA homeostasis, thereby inhibiting cholangiocyte proliferation. Therefore, JNK deficiency will stimulate the proliferation of cholangiocarcinoma cells and promote the development of cholangiocarcinoma. Increased proliferation is caused by changes in BA metabolism and increased liver expression of FXR / FGF15 / FGFR4, which triggers ERK activation in cholangiocellular cells, which in turn activates downstream proliferation-related genes. The activation of FXR by BA triggers human fibroblast growth Factor 15/19 (FGF15 / FGF19) secretion. Clinical studies have found that FGF15/19 is expressed in the liver during the development of hepatocellular carcinoma and intrahepatic cholangiocarcinoma. It is worth noting that FGF15/19 improves blood glucose response and reduces hepatic steatosis, while also promoting liver cancer. Therefore, the steady state change of BA causes FGF15/FGF19 to promote the development of cholangiocarcinoma.
In this process, liver PPARα is an important mediator of this regulatory cascade. PPARα deficiency significantly inhibits the phenotype caused by JNK deficiency. However, the role of PPARα in liver cancer remains unclear. Although some studies indicate that PPARα activation may promote liver cancer, other studies indicate that PPARα activation may be neutral or inhibit the development of liver cancer. This may be due to the different experimental conditions used in these studies. Recent studies have shown that the oncogenic effect of PPARα activation is partly due to changes in BA metabolism that drives ERK activation, which indicates that PPARα activation is a key factor in the development and progression of cholangiocarcinoma. The role of JNK /PPARα/ FGF signaling in lipid metabolism suggests that this pathway may represent a therapeutic target for steatosis and obesity. However, the potential risk of this pathway for carcinogenic progression indicates a serious problem with long-term treatment. Therefore, in the clinical treatment of steatosis and obesity using JNK inhibitory treatment strategies should consider the potential risk of cholangiocarcinoma.