Noroviruses are a frequent cause of both acute gastroenteritis and outbreaks of gastroenteritis. Its susceptible groups are usually children and the elderly. Norovirus is transmitted via the fecal-oral route, with different modes of transmission possible: person to person, via ingestion of food and/or water contaminated at source, food contaminated by food-handlers and through environmental contamination[1]. In low- and middle-income countries, the impact of norovirus infections in children may be more severe. According to statistics, there were approximately 200,000 children die each year from norovirus gastroenteritis[1, 2].
Specific diagnosis of norovirus is routinely made by polymerase chain reaction (PCR) assays or quantitative PCR assays, which give results within a few hours. These assays are very sensitive and can detect as few as 10 virus particles. A number of enzyme immunoassays (EIAs) are commercially available for the detection of norovirus GI and GII antigens in stool specimens. Compared to nucleic acid amplification tests, EIAs are generally simple to perform, do not require special molecular diagnostic laboratory facilities, and typically have a short turnaround time [3, 4].
Noroviruses encode a ~7.5 kb positive-sense, single-stranded RNA genome with three open reading frames (ORFs), which encode both the structural and the non-structural genes (Fig. 1) [5]. ORF1 encodes a ~200 kDa polyprotein, which is autoprocessed by a virally encoded protease to yield the non-structural viral replicase proteins essential for viral replication (Fig. 1) [5]. ORF2 encodes the 57 kDa major structural capsid protein VP1. ORF3 encodes a 22 kDa minor basic structural protein that has been hypothesized to function in packaging the genome into virions (Fig. 1). Norovirus protein functions are shown in Table 1 [6].
The Norovirus genus contains more than 40 different virus strains, which are divided into five genogroups based on sequence similarity. Genogroup I (GI), GII, and GIV viruses are primarily human pathogens, but there is a porcinespecific virus within the GII genogroup. GIII and GV viruses infect bovine and murine species, respectively. Each genogroup is further subdivided into genoclusters, based on sequence similarity and phylogenetic analysis. Comparison of the fulllength capsid sequence demonstrated that the five different genogroups varied by up to 60%, up to 57% among the human NoVs. In addition, sequences within a genocluster can vary as much as 20–30% in the capsid gene [5].
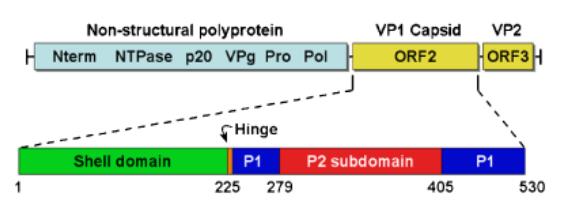 Fig. 1. The norovirus genomic structure and capsid domains.
Fig. 1. The norovirus genomic structure and capsid domains.
Table 1 Noroviruses proteins and their functions
*These functions have been proposed, but have yet to be demonstrated fully.
| Murine norovirus | Human noroviruses | Function |
| NS1/2 | p48 (N-term) | Replication complex formation*, contributes to persistence in MNV infections |
| NS3 | NTPase (2C-like) | RNA helicase*/NTPase |
| NS4 | p22 (3A-like) | Replication complex formation* |
| NS5 | VPg | Genome-linked protein involved in translation and replication |
| NS6 | Pro (3C-like) | Protease |
| NS7 | Pol/3Dpol | RdRp |
| VP1 | VP1 | Major capsid protein |
| VP2 | VP2 | Minor capsid protein |
| VF1 | No equivalent | Virulence factor |
First, the interaction of noroviruses with the cell surfis known to involve carbohydrate structures, which in the case of HuNVs include the histo blood group antigens (HBGAs). MNV binds host cells via sialic acid moieties, glycolipids and glycoproteins in a strain-dependent manner. But this is not sufficient to mediate entry and binding to an unidentified protein receptor is thought to be required. Then, the viral entry and uncoating phases proceed through as-yet-undefined pathways. The virus then turns on the synthesis of genetic material and the expression of proteins. The incoming viral genome is translated, through interactions with VPg (NS5) at the 59 end of the genome (red triangle in Fig 2) [5, 6] and the cellular translation machinery. The ORF1 polyprotein is co- and post-translationally cleaved by the viral protease NS6. The replication complex is formed by recruitment of cellular membranes to the perinuclear region of the cell (not shown), through interactions in part with NS1/2 and NS4. Genome replication occurs via a negative-strand intermediate, and genomic and subgenomic RNA are generated by the viral RdRp (NS7), using both de novo and VPg-dependent mechanisms of RNA synthesis. The replicated genomes are translated (within the replication complex) or packaged into the capsid, VP1, for virion assembly and exit. (Fig 2) [6].
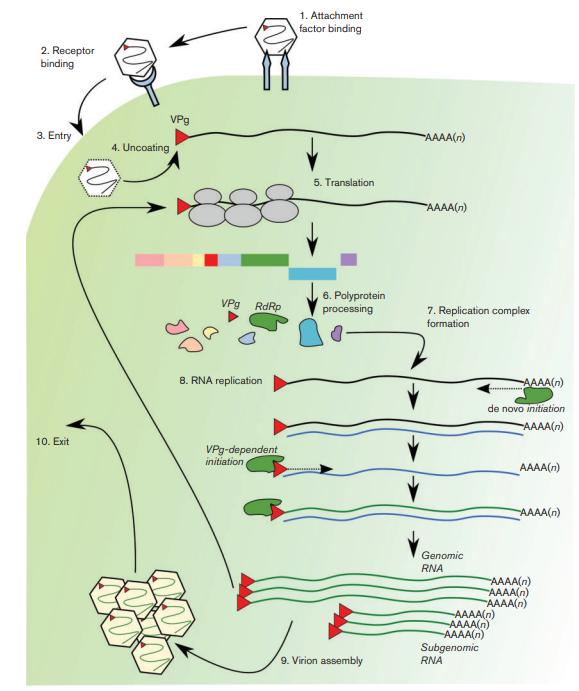 Fig 2. Outline of the norovirus life cycle.
Fig 2. Outline of the norovirus life cycle.
Vaccines against norovirus diseases have been shown to be of high significance. Among the vaccine candidates that are currently under clinical studies, recombinant VP1-based virus-like particles (VLPs) that mimic major antigenic features of noroviruses are the common ones, with proven safety, immunogenicity, and protective efficacy, supporting a high success likelihood of a useful norovirus vaccine [7,8].
The TAK-214 is an adjuvanted VLP-based bivalent vaccine containing two types of VLPs, one from the GI.1 Norwalk virus (NV) strain and the other one from a consensus GII.4 sequence (GII.4c) derived from three GII.4 variants. Several phase 1 clinical studies were conducted, proving good safety and tolerability. In a phase 2a trial, two doses of intranasally delivered vaccine using chitosan and monophosphoryl lipid A (MPLA) as adjuvants were demonstrated to protect healthy adults against challenge of homologous GI.1 NV. The first in-human phase 2b field efficacy study of the TAK-214 vaccine was conducted over two winter seasons in a U.S. Navy training facility with over 4600 participants. the vaccine efficacy against GII.2 noroviruses was 57.4% (95% CI, 7.0 to 80.5; p = 0.0321), indicating that the TAK-214 vaccine elicits a broad immune response and confers cross-genotype protection [9, 10].
Another norovirus vaccine candidate in clinical study is a recombinant VP1-based bivalent vaccine (VXA-NVV-104). This vaccine contains recombinant adenovirus-based vectors [11] carrying genes encoding noroviral VP1s to express VP1 antigens locally in the epithelial cells within the intestine of vaccine recipients to induce mucosal immunity. The adenovirus vector also contains specific RNA-encoding sequences to generate double-stranded RNAs as an adjuvant for enhanced immunogenicity of the VP1 antigens. Both VP1-encoding genes of the GI.1 NV strain and the GII.4 Sydney variant are included as a bivalent vaccine for broad efficacy. Therefore, this tablet vaccine is designed to be administered orally to elicit the mucosal immune response locally within the intestine to prevent norovirus infection [8, 11].
The third norovirus vaccine candidate under clinical study is a bivalent VLP-based vaccine (Hansenulapolymorpha) that was developed by the National Vaccine and Serum Institute (NVSI) of China. It is composed of two recombinant VLPs representing the GI.1 and the GII.4 genotypes, respectively, with aluminum salt as the adjuvant [8]. And the forth norovirus vaccine is the Longkoma Norovirus Vaccine. This is a quadrivalent vaccine consisting of four aluminum salt adjuvanted VP1 proteins, each representing GI.1, GII.3, GII.4, or GII.17 genotypes, produced by means of a yeast expression system [8].
Noroviruses are the major viral pathogens causing epidemic and endemic acute gastroenteritis with significant morbidity and mortality. The development of a broadly effective norovirus vaccine remains difficult, owing to the wide genetic and antigenic diversity of noroviruses with multiple co-circulated variants of various genotypes [8, 9]. In addition, the absence of a robust cell culture system, an efficient animal model, and reliable immune markers of norovirus protection for vaccine evaluation further hinders the developmental process. Noroviruses are known for their wide genetic and antigenic diversity. Infection by a single norovirus strain does not appear to confer a broad, long-lasting immunity. An even more complex panel of antigens representing more norovirus genotypes should be considered for even broader vaccine efficacy. In addition, the adjuvant is an important component of a successful vaccine to enhance the magnitude, breadth, and durability of the vaccine antigens. The current success of the mRNA vaccine platform in development of COVID19 vaccines, as well as its further applications in developing vaccine candidates against influenza and malaria, provide a new approach to advance the norovirus vaccine development program [8, 11].
References
| Target | Capture | Detection | Application |
| Norovirus | DMAB4016 | DMAB4017 | ELISA |
| DCABY-4778 | DCABY-4779 | ELISA | |
| CABT-Z670M | CABT-Z671M | ELISA, LFIA | |
| CABT-Z671M | CABT-CS1031 | ELISA, LFIA | |
| Norovirus GI Capsid | CABT-B8765 | CABT-B8766 | ELISA |
| CABT-CS1032 | CABT-B8765 | LFIA | |
| DMABP-L27 | DMABP-L26 | ELISA, LFIA | |
| Norovirus GII | CABT-B8767 | CABT-B8768 | ELISA |
| DCABY-4585 (GII.4) | DCABY-4586 (GII.4) | ELISA | |
| CABT-CS1036 | CABT-B8768 | LFIA | |
| CABT-CS1038 (GII.2) | CABT-CS1039 (GII.4) | ELISA, LFIA | |
| DMABP-L28 | DMABP-L29 | ELISA, LFIA |
| Cat | Product Name | Host | Application | |
| CABT-NS1353 | Anti-Norovirus GII.4 P Domain Mab | Human | Neut, BL, ELISA | Inquiry |
| CABT-NS1354 | Anti-Norovirus GII.4 VP1 Mab | Human | Neut, ELISA | Inquiry |
| CABT-NS1355 | Anti-Norovirus GII.4 VP1 Mab | Mouse | Neut, ELISA | Inquiry |
| CABT-NS1357 | Anti-Norovirus GII.4 P Domain Mab | Mouse | Inhib | Inquiry |
| CABT-YN1228 | Anti-Murine Norovirus-1 Capsid P Domain Mab | Human | Neut, ELISA, BL | Inquiry |
| CABT-YN1234 | Anti-Norovirus GII.4 Mab | Human | Neut, Inhib | Inquiry |
| CABT-YN1305 | Anti-Murine Norovirus-1 Capsid P Domain Mab | Mouse | Neut, ELISA, BL | Inquiry |
| CABT-YN1311 | Anti-Norovirus GII.4 Mab | Mouse | Neut, Inhib | Inquiry |
| Cat. | Product Name | Species Reactivity | Sample | |
| DEIANS084 | Mouse Anti-Norovirus (MNV1/VP1) IgG ELISA Kit, Quantitative | Mouse | Serum, plasma or other biological fluids. | Inquiry |
| DEIANS085 | Anti-Norovirus GI.1 Virus ELISA Kit, Quantitative | Human | Serum and Plasma | Inquiry |
| DEIANS086 | Human Anti-Norovirus GI.1 VP1 IgM ELISA Kit, Quantitative | Human | Serum and Plasma | Inquiry |
| DEIANS087 | Monkey Anti-Norovirus GI.1 VP1 IgG ELISA Kit, Quantitative | Monkey | Serum and Plasma | Inquiry |
| DEIANS088 | Anti-Norovirus GII.4 Virus ELISA Kit, Quantitative | Human | Serum and Plasma | Inquiry |
 Fig. 1 Reactivity of Norovirus Mab sets
Fig. 1 Reactivity of Norovirus Mab sets
a

b
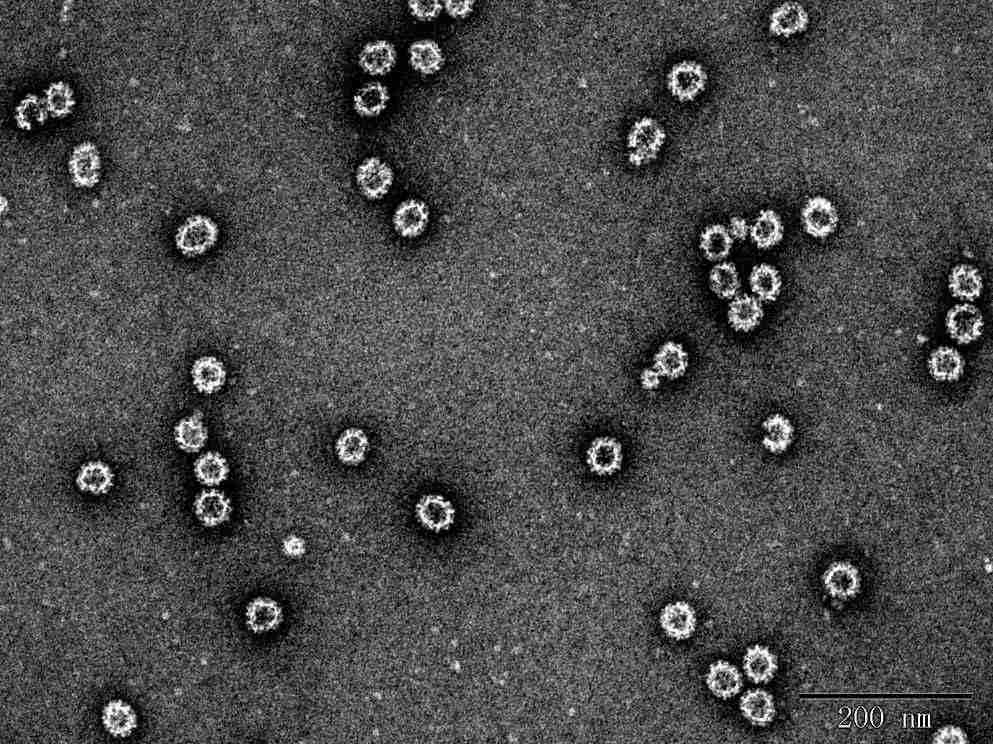
c
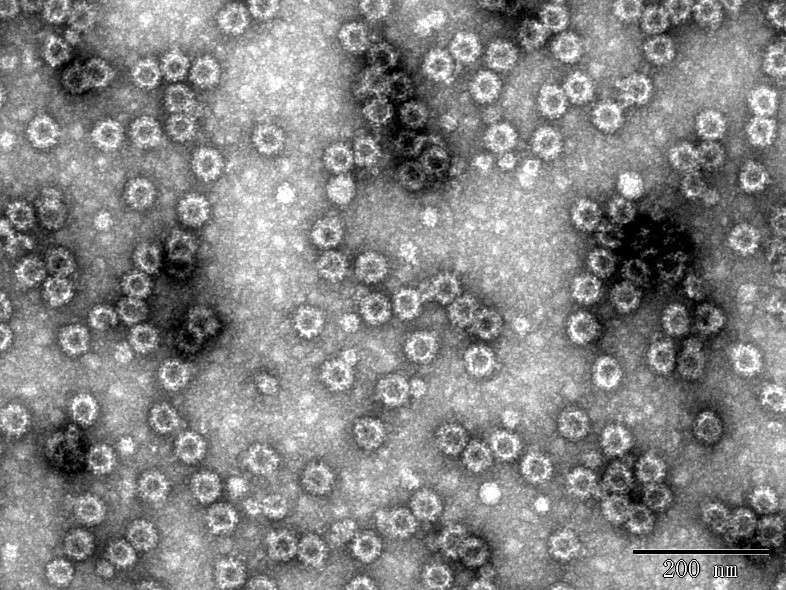
d
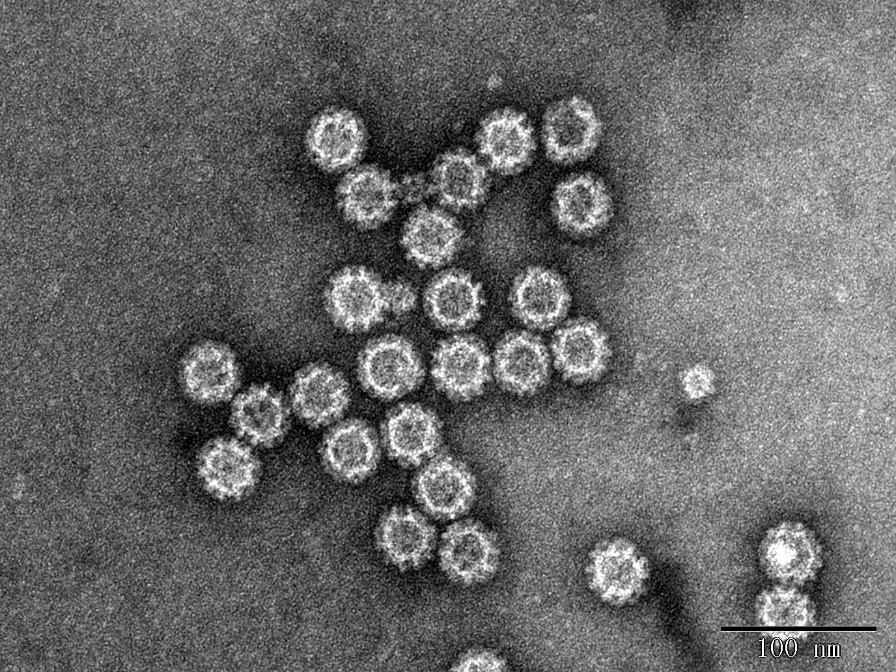
e

f
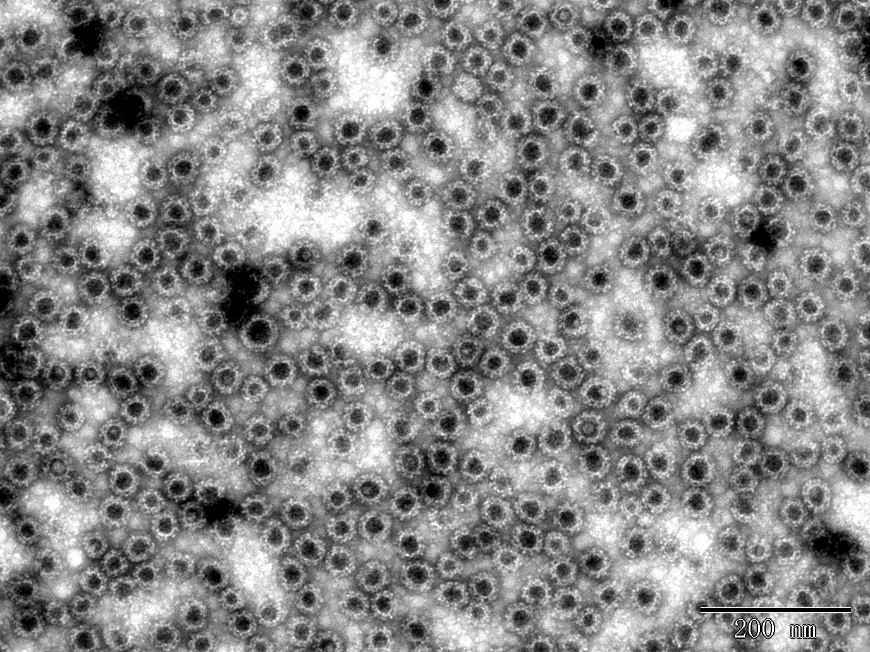
g

Fig. 2 TEM images of Norovirus VLP
a. NoV GI.1 VLP (DAG-WT2451).
d. NoV GII.4 VLP (DAG-WT2454).
g. NoV GII.17 VLP (DAG-WT2457)
b. NoV GII.2 VLP (DAG-WT2452).
e. NoV GII.6 VLP (DAG-WT2455).
c. NoV GII.3 VLP (DAG-WT2453)
f. NoV GII.13 VLP (DAG-WT2456)
