
The structure of all immunoglobulins is basically the same, consisting of two heavy chains (Heavy, abbreviated as H) and two light chains (Light, abbreviated as L), and looks like a “Y”-shaped symmetrical macromolecule. The stem of “Y” is called the Fc segment, which consists of the -COOH group end half of the two heavy chain polypeptide chains. The F c segments of the two heavy chains are connected by one or several disulfide bonds. The two forks of “Y” are called F ab segments, and each fork is composed of a whole light chain and half of the -NH base end of the heavy chain. The distal end of each fork has an antigen binding site with the same characteristics. This structural model has been confirmed by electron micrographs. At the same time, it was also confirmed that the Fc segment and two Fab segments of a complete antibody molecule are connected by a hinge region. There are a lot of proline in the hinge area, the structure is extremely loose, and it has the characteristics of toughness and flexibility. It can adjust the angle between Fab segments to adapt to the spatial changes between antigenic determinants, so as to coat antigens more effectively.
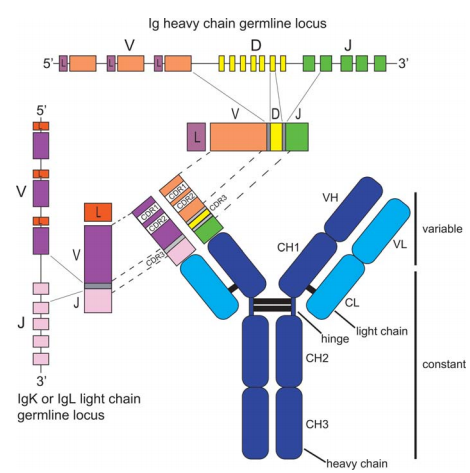
For the same type of antibody, the most characteristic feature that constitutes each immunoglobulin molecule is expressed in the 110-120 residues of the -NH base end of the polypeptide chain. The amino acid sequence of this part is different from antibody protein to antibody protein. However, the rest of the antibody polypeptide chain is constant. Therefore, according to the amino acid sequence of the constant region, the light chain can be divided into two types, namely K (kappa) and λ (Lamda). Among them, K has no subtype, while the constant region of the λ type has a certain change. By analyzing the amino acid sequence at positions 1-20 of -NH2, it can be typed again. Among them, K is divided into I, II, and III, and λ is divided into I, H, I, W, and V. The K subgroup cannot be combined with the lambda type, and vice versa. Further research found that a plasma cell can only produce one type.
The heavy chain can also be classified according to the amino acid sequence of its constant region, which is divided into five categories: IgG (γ chain), IgM (μ chain), IgA (α chain), IgD (δ chain) and IgE (ε chain). Similar Ig Molecules are divided into several subclasses according to the position and number of disulfide bonds between chains. For example, IgG is divided into four subclasses, IgM is divided into two subclasses, and IgA is divided into two subclasses. There are 4 disulfide bonds in the γ and α chains, and 5 (or more) disulfide bonds in the μ, ε and δ chains. According to the characteristics of 1-20 amino acids at the base end of -NH, the heavy chain can be divided into four subgroups. The subgroups of the H chain are different from the subgroups of the L chain, and they can be combined with the constant regions of other heavy chains. For example, VHI can be combined with the constant region of α and γ.
The above classes, subtypes and types, subtypes and subgroups are all of the same genus, and the Ig specificity shared by all individuals is called isotype. The same species but different individuals, the polypeptide chain can have one or more amino acid differences, which is called allotype. This situation is very similar to the situation of ABO blood type and major histocompatibility antigen (H-LA). Generally speaking, Gm is the type of IgG. There are 30 such genotypes. The Gm factor is located at CH1-3. There is also the Am type of IgA, which is mainly determined by the α subtype, namely Am1-2. The difference lies in the presence or absence of disulfide bonds. Km is mainly different from the amino acid at position 191. For example, when it is leucine, it is called Km1; when it is valine, it is called Km3. Except the genotype of IgG is determined by the Fc segment, everything else is determined by the Fd segment. In addition, there is a so-called idiotype, which is determined by the antibody-producing cell. In the same genus, each individual produces different antibodies when stimulated by the same antigen, so there are countless idiotypes. Another notable feature of the polypeptide chains of the H and L chains is that each chain includes a homologous segment of approximately 110 residues. Its characteristics are as follows:
1) These fragments show homology in the amino acid sequence of either the H chain or the L chain constant region. CL and CH, CH2, and CH are similar in structure, that is, they have homology.
2) These fragments contain an intrachain disulfide bond between the two cysteines, which is separated by approximately 65 amino acids.
3) Each fragment is folded independently of other fragments to form a globular protein.
4) The spherical structure of different chains is a unit with unique functions generated by the interaction of non-covalent bonds. When this fragment is separated from the entire molecule, its function is naturally retained. Different functional areas have different functions, they can bind antigen, complement, or cells. Therefore, after the antibody molecule is divided by various enzymes, fragments of Fab, F(ab)2, Fc, Fabc or Fv can be produced.
secondly, antibody is the existence of hypervariable areas. As mentioned earlier, different antibodies have different variable region sequences. Both the H chain and the L chain have a special amino acid sequence region, which is more variable than the rest of the V region. These regions are called hypervariable regions of variable regions. Variability is determined by the ratio of the frequency of different amino acids at a certain position to the most common amino acid at that position. Of the 110 variable region amino acids in the light chain, 25 are hypervariable. 30 of the 120 amino acids in the heavy chain are hypervariable. There are three light chains and three heavy chains. The light chain hypervariable region and the heavy chain hypervariable region are different. Recognizing one antigenic determinant requires a set of heavy chain and light chain hypervariable regions to complement it, and another antigenic determinant requires another set. Complementary hypervariable regions. At present, more than 200 kinds of hypervariable regions have been discovered, and their different sets can recognize countless antigenic determinants. The relatively constant region between hypervariable regions is called the framework region. Because the variability of the amino acid sequence outside the hypervariable region is very small, this indicates that the hypervariable region alone forms a highly specialized surface for the binding site. This has been confirmed by different hapten-labeled affinity experiments. The amino acids in the framework region also have some mutation rates, but they are very low. Therefore, it can be divided into subgroups accordingly. The main function of the framework region is to facilitate the movement of the hypervariable region in three-dimensional space, such as forming a cavity or shallow groove to fix the antigen.
Antigen-antibody binding through non-covalent bonds is reversible but specific. Moreover, it was further discovered that not all hypervariable regions are involved in binding antigen, which is related to the number of antigenic determinants. And it proves that the role of H is greater than that of L, but they work together.
The so-called affinity (affinity) refers to the binding ability of antibody molecule Fab and antigen, and avidity refers to the ability of the entire antibody molecule to bind to the antigen. The two are different. Because IgM is a pentamer, the avidity of IgM is stronger than IgG, but their affinities are equal. It has been confirmed that the functional regions of all immunoglobulins show a very similar three-dimensional structure. In this folding, the three-dimensional size of a functional area is 40 x 25x25A, composed of two layers, which are connected by disulfide bonds. There are many non-covalent bonds between the two layers, which play a stabilizing role.
These hierarchical filamentous structures are connected by loops of variable length, and hypervariable region residues are located on some loops. Immunoglobulins are produced by B cells. The average concentration of various types of immunoglobulin in normal human serum and the concentration outside the blood vessel are different. IgG is the most common protein in serum, and it is most easily transferred through the cell membrane, so its distribution in blood vessels and outside blood vessels is almost equal. One of the main functions of IgG is to neutralize soluble antigens, such as bacterial toxins. IgM exists in blood vessels and is mainly responsible for processing particulate antigens in serum. lgM has a high agglutination effect and the role of complement binding. IgA can be present in serum, but more importantly in secretions (saliva, colostrum, tears, nose, trachea, and intestinal secretions). In these secretions, Ig A forms a barrier against microorganisms invading and mucosal surfaces. IgE is present in the serum and is distributed in the exocrine fluid at a very low concentration. It is mainly related to the body’s allergic reaction. It can connect to mast cell receptors and bind to allergens, causing the release of histamine. IgD is the only immunoglobulin that can be hydrolyzed by plasma proteases. It is produced in the plasma cells of the tonsils and other glands.